De Grutto (Limosa limosa) in de Kalkense Meersen.
Historiek, broedsucces en toekomstperspectieven
Robbert Schepers

foto : Marc Van den Bril
Scriptie voorgelegd tot het behalen van de graad van Master in de Biologie
Academiejaar 2009/2010
Promotor: Prof. dr. Johan Mertens
Begeleiders: Dr. Jenny De Laet, ABLLO vzw
Peter Claus, Regionaal Landschap Schelde-Durme
1 Inleiding
1.1 Algemeen
Bijna alle broedende weidevogels in Europa vertonen de laatste decennia een afname in populatiegrootte (Thorup 2006). Een daling in het broedsucces wordt aangeduid als de drijvende kracht achter deze trend (Green 1988, Peach et al. 1994, Besbeas et al. 2002, Ottvall 2005, Schekkerman et al. 2008). Men neemt aan dat de intensivering van het agrarisch graslandgebruik de voornaamste oorzaak is van deze achteruitgang (Beintema et al. 1997, Kruk et al. 1997, Vickery et al. 2001, Wilson et al. 2004, Schekkerman & Beintema 2007).
Het landgebruik op grasland is in de tweede helft van de twintigste eeuw aanzienlijk veranderd en een groot deel van de laag gelegen graslanden wordt tegenwoordig intensief bewerkt. De belangrijkste gevolgen van deze intensivering van de landbouw zijn een verlaging van de grondwatertafel, een verdubbeling van de mestgift, een omschakeling van hooi naar ingekuild gras en hogere veedichtheden. Structuur- en soortenrijke grasvegetaties werden vervangen door dichte, snelgroeiende, uniforme grasmatten die gedomineerd worden door competitieve soorten. Deze ommekeer heeft ertoe geleid dat grasland tegenwoordig minder geschikt is als foerageer- en broedhabitat voor vogels (Vickery et al. 2001).
Bemesting en het verlagen van de grondwatertafel bevorderen een snellere groei van het gras waardoor er steeds vroeger kan gemaaid worden. De vogels passen zich wel enigszins aan, maar ze kunnen deze wedren toch niet helemaal bijhouden met als gevolg dat heel wat nesten en jongen verloren gaan door uitmaaien (Beintema et al. 1995). Gemaaid grasland biedt minder bescherming voor de kuikens waardoor ze gemakkelijker gepredeerd worden (Schekkerman et al. 2005). Reeds gemaaid grasland is bovendien niet geschikt voor gruttokuikens die foerageren op in de vegetatie levende insecten. Op gemaaide percelen zijn namelijk maar half zoveel insecten aanwezig in vergelijking met ongemaaide. Om deze reden vinden kuikens niet veel voedsel en moeten ze veel meer tijd besteden aan foerageren (Schekkerman et al. 1997). De vegetatie op bemeste percelen is meer uniform en de structurele diversiteit die gevonden wordt op niet-bemeste percelen ontbreekt. De nesten van weidevogelsoorten die berusten op cryptisch gekleurde eieren, zoals grutto (Limosa limosa) en kievit (Vanellus vanellus), zijn hierdoor meer zichtbaar hetgeen predatie in de hand werkt (Baines 1990). Bovendien is er de directe invloed van de mestaanwending op de nesten met eieren. Nieuwe landbouwtechnieken zoals mestinjectie reduceren de kans dat een nest overleeft tot een minimum en de toegenomen veedichtheden resulteren in grote verliezen door vertrapping (Beintema et al. 1995).
Intensieve bemesting heeft eveneens een invloed op de voedselbeschikbaarheid. Bij toenemende bemesting daalt de kruidenrijkdom van de vegetatie en als gevolg de diversiteit aan insecten waarmee kievit- en gruttojongen zich voeden (Beintema et al. 1995). Siepel (1990) constateerde ook een sterke afname van de gemiddelde grootte van de vegetatiefauna bij toenemende bemesting. De abundantie van regenwormen, samen met emelten het stapelvoedsel van volwassen grutto’s, neemt bij bemesting daarentegen toe (o.a. Standen 1984, Hansen & Engelstad 1999).
Bijgevolg trappen grutto’s en andere weidevogels in een ‘ecologische val’. Ze kiezen een nestplaats op bemeste percelen omdat daar voor volwassen individuen meer voedsel aanwezig is (Kleijn et al. 2001) en omdat het gras daar langer is, wat ze prefereren om hun nest te maken (Beintema et al. 1995). De bemeste percelen beschikken echter over een lager voedselaanbod voor de jongen en worden sneller gemaaid dan onbemeste reservaatpercelen. Voor de kievit geldt een gelijkaardig scenario, maar dan vooral op maïsland waar de verliezen hoofdzakelijk een gevolg zijn van het ploegen, rollen en inzaaien van de gewassen.
Intensivering van de landbouw is echter niet de enige oorzaak van de achteruitgang. Recent wordt ook het belang van toenemende predatiedruk benadrukt (Teunissen et al. 2008). Predatie is de meest voorkomende natuurlijke verliesoorzaak bij weidevogels (Bos & Vugteveen 2005). Meestal worden zwarte kraaien en de vos door natuurbeschermers en landbouwers aangeduid als de belangrijkste predatoren. Er is echter nog weinig geweten over de identiteit van de predatoren die verantwoordelijk zijn voor het roven van eieren of het doden van jongen.
Een studie uit Nederland, waar nesten werden uitgerust met camera’s en temperatuursensoren, trachtte hierop een antwoord te formuleren (Teunissen et al. 2008). De auteurs troffen een grote variatie aan in de identiteit van predatoren. Eieren werden vooral door zoogdieren gepredeerd en kuikens door vogels. Onder de predatie van eieren waren de vos (Vulpes vulpes) en de hermelijn (Mustela erminea) de belangrijkste zoogdieren en in mindere mate ook andere marterachtigen. Kuikens werden vooral geroofd door buizerd (Buteo buteo), blauwe reiger (Ardea cinerea), hermelijn en in mindere mate zwarte kraaien (Corvus corone). Bos & Vugteveen (2005) vermelden dat ratten ook belangrijkere predators kunnen zijn dan wordt gedacht.
In Nederland, waar ongeveer de helft van de (noord)West-Europese gruttopopulatie broedt (Hagemeijer & Blair 1997), proberen de overheid en natuurbeschermingsorganisaties al lange tijd weidevogels te beschermen via agrarisch natuurbeheer. Dit gebeurt in reservaten maar ook door het afsluiten van contracten met boeren, die een financiële tegemoetkoming krijgen voor het later maaien of beweiden van percelen, het beschermen van nesten, enz. Sommige maatregelen zijn toegesneden op de grutto maar men neemt aan dat deze ook voor andere weidevogelsoorten gunstige effecten hebben. Recente studies tonen aan dat deze beheerovereenkomsten echter niet het gewenste resultaat opleveren (Kleijn et al. 2001, Schekkerman et al. 2005). De gegevens van Schekkerman et al. (2005) wekken zelfs de indruk van een teruglopend broedsucces van Nederlandse grutto’s in de afgelopen decennia, ook in gebieden waar volgens de geldende inzichten voldoende kuikenland (voor jongen geschikt foerageerhabitat, m.a.w. lang en niet gemaaid gras) aanwezig was. Maatregelen zoals een uitgestelde maaidatum kunnen wel een gunstige invloed hebben op de overleving van nesten en kuikens, maar het reproductief succes van grutto’s is gemiddeld onvoldoende om de jaarlijkse sterfte van volgroeide vogels te compenseren (Schekkerman & Müskens 2000). Als er al sprake is van een hoger broedsucces in gebieden met agrarisch natuurbeheer is dit meestal te wijten aan een hogere uitkomstkans van legsels, die op zijn beurt weer het gevolg is van geringere agrarische nestverliezen. De mortaliteit van kuikens ligt daarentegen meestal nog te hoog. Aangezien de overleving van kuikens een positieve relatie vertoont met het gemiddelde aanbod aan niet gemaaide percelen wijzen maatregelen om het agrarisch natuurbeheer effectiever te maken in de richting van een hoger aanbod aan niet gemaaid grasland tijdens de belangrijkste kuikenperiode (Schekkerman et al. 2005).
Ondanks de negatieve vaststellingen in Nederland en andere landen inzake populatiegrootte (Trolliet 2000, Altenburg & Wymenga 2000, Teunissen 2000, Busche 1994), wordt deze negatieve trend niet vastgesteld in België. Volgens Devos et al. (2003) is er zelfs sprake van een toename van het broedbestand bij kievit en grutto en ook Vermeersch et al. (2006) stellen een positieve trend vast. Vermeersch et al. (2006) leggen wel de nadruk op een meer kritische en terughoudende beoordeling van de toestand van de weidevogels in Vlaanderen aangezien de positieve trends die zij observeerden ten dele kunnen vertekend zijn door de erg gunstige weersomstandigheden in de waarnemingsperiode. In de kerngebieden blijkt het broedsucces van grutto’s echter erg laag te zijn, zoals blijkt uit een studie in het Antwerpse Linkeroevergebied. In de periode 1997-2002 overleefden daar slechts 0,18-0,26 gruttojongen per broedpaar, alhoewel voor de instandhouding van een populatie 0,5-0,8 vliegvlugge jongen per paar vereist zijn. (Van Impe 2004, Schekkerman & Müskens 2000). Ook voor de kievit was het broedsucces te laag.
Er zouden dus aanwijzingen zijn dat de toename van de Vlaamse weidevogelpopulaties eerder te verklaren is door nieuwe vestigingen van adulte vogels van elders dan door effectieve jongenaanwas uit de lokale populaties.
De Vlaamse populaties fungeren dan mogelijk als sink-populatie (Vermeersch et al. 2006).
Ook in het studiegebied van dit onderzoek, de Kalkense Meersen, steeg de populatie grutto’s de afgelopen jaren,
met een maximum van 30 koppels in 2002. Momenteel stagneert de populatie rond 25 broedparen. Aangezien een specifiek weidevogelbeheer nodig is in het agrarische gebied om de weidevogelpopulaties in stand te houden (Beintema 1995, Schekkerman & Müskens 2000) en als voorbereiding op het geactualiseerde Sigmaplan (zie verder) is het Regionaal Landschap Schelde-Durme in opdracht van het Agentschap voor Natuur en Bos en in samenwerking met de Provincie Oost-Vlaanderen in 2008 gestart met een soortenbeschermingsproject rond weidevogels in de Kalkense Meersen. Dit weidevogelproject heeft twee doelstellingen. Enerzijds de instandhouding en uitbreiding van de broedpopulatie van weidevogels, grutto in het bijzonder, met een belangrijk accent op de Sigmagebieden van de cluster Kalkense Meersen. Anderzijds het ontwikkelen van samenwerkingsverbanden en netwerken tussen o.m. lokale landbouwers, natuurbeschermers en jagers, in het bijzonder aangaande weidevogels en weidevogelbescherming.
Op deze manier krijgen de landbouwers reeds ervaring met het weidevogelbeheer dat opgelegd zal worden bij de uitvoering van het geactualiseerde Sigmaplan. Het project wil op perceelsniveau invulling geven aan de bescherming van weidevogels door de landbouwer, deels tegen vergoeding, enkele pakketten maatregelen aan te bieden die het succesvol grootbrengen van een broedsel bevorderen. De beschikbare pakketten zijn :
Rustperiode tussen 1 april en 15 juni (geen landbewerkingen): vergoeding van 350 euro per ha.
Rustperiode tussen 1 april en 1 juni (geen landbewerkingen): vergoeding van 200 euro per ha.
Extensieve beweiding tussen 1 april en 1 juni (2 runderen per ha): vergoeding van 250 euro per ha.
Bescherming van individuele nesten door vrijwilligers: geen vergoeding.
In 2008 en 2009 besloegen de beheersovereenkomsten een oppervlakte van respectievelijk 45 ha en 68 ha. In 2010 steeg de oppervlakte onder beheersovereenkomsten verder tot ongeveer 90 ha.
In zo’n project is er uiteraard nood aan monitoring om na te gaan of de getroffen maatregelen wel degelijk tot resultaten leiden. In het verleden zijn er reeds inventarisaties uitgevoerd van het aantal broedparen, maar er werd nog nooit enige vorm van onderzoek gedaan naar het broedsucces van de weidevogels (persoonlijke mededeling Peter Claus). In het kader van het huidige weidevogelproject werd een dergelijk onderzoek gevraagd door het Agentschap voor Natuur en Bos. Deze vraag vormt het onderwerp van deze studie, met een focus op de grutto.
1.2 Het geactualiseerd Sigmaplan
Als reactie op de stormvloed van 1976 werd in 1977 het Sigmaplan opgesteld en in uitvoering gebracht. Dit plan moest het getijgebonden gedeelte van de Schelde en haar zijrivieren in Vlaanderen beveiligen tegen overstromingen als gevolg van stormvloeden uit de Noordzee. Het Sigmaplan is lange tijd een plan geweest met als enige doelstelling het bereiken van voldoende veiligheid. Het plan is echter nooit volledig afgewerkt. Bovendien zou het, zelfs als het nu afgewerkt zou zijn, niet meer de veiligheid kunnen garanderen waarvoor het ooit ontworpen werd omdat de fysische omstandigheden intussen gewijzigd zijn. Stormvloeden komen frequenter voor en de waterpeilen zijn hoger. Daarenboven wordt nog een verdere zeespiegelstijging verwacht als gevolg van de klimaatopwarming. Omwille van bovenstaande redenen en als gevolg van vooruitstrevende inzichten op vlak van multifuntionaliteit van rivierecosystemen was er nood aan een nieuw plan. Het geactualiseerde Sigmaplan bevat naast veiligheid ook de pijler ‘natuurlijkheid’. Een aanzienlijk deel van het Schelde-estuarium is immers van groot ecologisch belang (Natura 2000). Door het opnemen en creëren van natuurkerngebieden, in combinatie met veiligheid, moet het Sigmaplan bijdragen tot het behalen van de instandhoudingsdoelstellingen (IHD).
Het optimale Sigmaplan (veiligheid) en het optimale natuurontwikkelingsplan werden geïntegreerd in het Meest Wenselijke Alternatief (MWeA) nadat een consensus ontstond met de belangengroepen. Het MWeA is het uitgangspunt voor de concretisering van het geactualiseerde Sigmaplan. In het MWeA worden habitatdoelstellingen vooropgesteld om de IHD per deelgebied te halen. Deze habitatdoelstellingen geven een invulling aan de systeem- en soortdoelstellingen.
De Kalkense Meersen vormen een deelgebied van het Sigmaplan en conform het MWeA zal het vanaf 2010 volledig ingericht worden als ‘wetland’ waarin o.a. plaats is voor een weidevogelgebied van 180-200 ha (zie verder, 3.1.1).
Het weidevogelproject kadert in dit geactualiseerde Sigmaplan en dient om de tussentijdse periode tot aan de inrichting van het weidevogelgebied te overbruggen. Deze overgangsmaatregelen hebben als doel de weidevogelpopulatie (in het bijzonder de grutto) zo robuust mogelijk te houden, zodat bij aanvang van de realisaties i.k.v. het geactualiseerd Sigmaplan op het terrein gestart kan worden vanuit een zo optimaal mogelijke uitgangssituatie.
2 Doelstellingen
In het oorspronkelijk opzet streefde het onderzoek vier doelstellingen na:
1. Inventariseren van het aantal broedparen van grutto en kievit in de Kalkense Meersen en het Aubroek te Berlare.
2. Bepaling van het broedsucces van grutto’s in termen van bruto territoriaal succes (BTS) in:
Een gebied met hoofdzakelijk bemeste percelen zonder beheersovereenkomsten (Het Aubroek, Berlare).
Een gebied met hoofdzakelijk bemeste percelen met beheersovereenkomsten (Broekmeers, Kalkense Meersen).
Een gebied met hoofdzakelijk onbemeste percelen waar aan natuurbeheer gedaan wordt (‘Reservaatzone’, Kalkense Meersen).
3. Onderzoeken of het weidevogelproject effectief zorgt voor een hoger aandeel niet gemaaid grasland in de kuikenperiode.
4. Opstellen van een historisch overzicht van het aantal broedkoppels en de broedlocaties van de gruttopopulatie in de regio Kalkense Meersen.
Na het opstellen van het historisch overzicht bleek een duidelijke verschuiving van de broedlocaties naar bemeste percelen. Deze vaststelling riep nieuwe vragen op en als antwoord daarop werden nieuwe doelstellingen toegevoegd aan het onderzoek:
5. Bepaling van het aantal en de biomassa regenwormen aanwezig in de bodem van bemeste en onbemeste percelen.
6. Bepaling van de hoogte van de vegetatie in bemeste en onbemeste percelen
7. Bepaling van de kruidenrijkdom tussen bemeste en onbemeste percelen
Voor het onderzoek naar het BTS van de grutto’s werd een hoger BTS verwacht in gebieden onder beheer of beheersovereenkomsten omdat daar normaal gezien minder verliezen door landbouwwerkzaamheden optreden en omdat niet gemaaid gras meer voedsel en bescherming biedt aan kuikens.
De belangrijkste doelstelling van het weidevogelproject is het aanbieden van een hoger aandeel niet gemaaid grasland, t.o.v. onbeheerde percelen. De verwachtingen zijn dan ook dat dit effectief zo is.
Om effecten van het weidevogelproject op de populatiegrootte te evalueren en om bepaalde trends waar te nemen en te verklaren is het zinvol een historisch overzicht op te stellen van de gruttopopulatie. Het historisch verloop van de broedlocaties werd opgesteld naar aanleiding van een vermoeden dat er de laatste jaren een verschuiving plaatsvindt in broedlocaties van onbemeste naar bemeste percelen in het algemeen en van Molenmeers naar Broekmeers in het bijzonder. Deze verschuiving is mogelijk te verklaren doordat op de bemeste percelen (Broekmeers) meer voedsel aanwezig is onder de vorm van regenwormen en dat de vegetatie er hoger is op het moment dat de grutto’s arriveren en hun nestplaats kiezen.
Tenslotte werd de kruidenrijkdom tussen bemeste en onbemeste percelen onderzocht omdat dit belangrijke consequenties kan hebben voor de voedselbeschikbaarheid voor de jongen. Bemeste percelen zijn doorgaans minder divers en hebben als gevolg een minder rijke insectenfauna.

Luchtfoto van de Kalkense Meersen. (Ludo Goossens © Vilda)
3 Materiaal & methoden
3.1 Studiegebied
3.1.1 Kalkense Meersen
Het volledige gebied van de Kalkense Meersen is ongeveer 800 ha groot en ligt op het grondgebied van de gemeentes Wetteren, Laarne (Kalken), Wichelen (Schellebelle) en Berlare (Overmere, Uitbergen). Het wordt begrensd door de hoger gelegen (+5m TAW) dorpskernen van Overschelde, Heesvelde, Kalken, Overmere en Uitbergen en in het Zuiden door de Zeeschelde. In het noordoosten sluiten de Kalkense Meersen aan op het Donkmeer en Berlare Broek, een relatief uitgestrekt laagveengebied. De Kalkense Meersen bestaan voornamelijk uit vochtige graslanden en de belangrijkste toponiemen zijn Kastermeersen, Oude Schelde, Broekmeers, Molenmeers, Langendonk, Scherenmeersen, Belham, Heisbroek en Wijmeers (fig. 3.1).
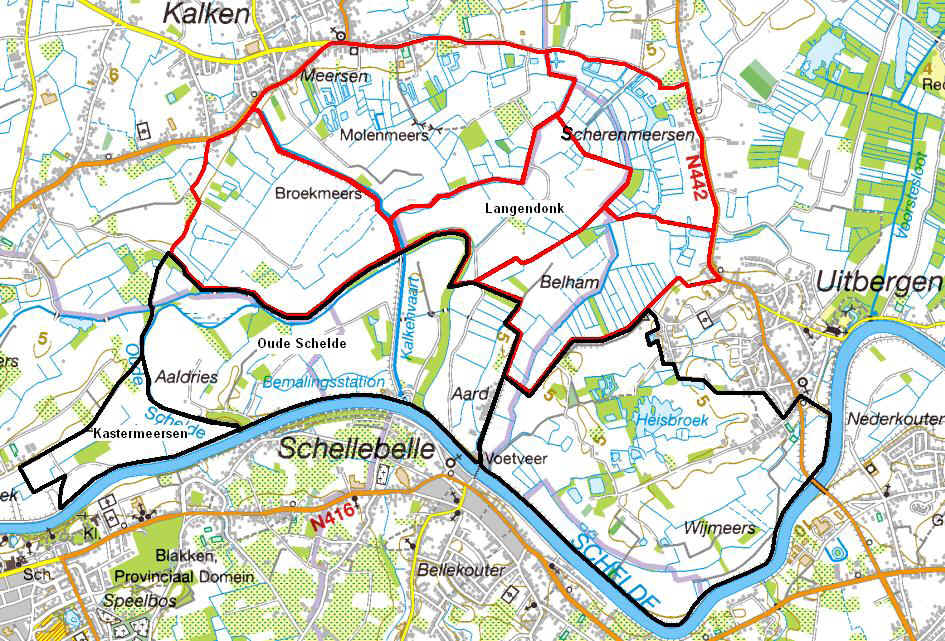
Figuur 3.1. Belangrijkste toponiemen in de Kalkense Meersen. Deelgebieden van het studiegebied van dit onderzoek zijn aangeduid in rood.
De Kalkense Meersen zijn gelegen in de laagvlakte van ‘De Vlaamse Vallei’ en vormen één van de grootste overblijfselen van de Scheldemeersen. Het is een alluviale vlakte die omstreeks de 12e eeuw ingepolderd werd en ligt in een oude meander van de Schelde (10000 jaar oud). Het gebied bestaat voornamelijk uit kleiige komgronden (natte klei en natte zware klei), ontstaan door overstromingen vanuit de rivier. Het zwaarste sediment wordt langs de oevers afgezet, waardoor zandige oeverwallen ontstaan. Het lichtere sediment bezinkt pas verder van de rivier in de komgronden. Langs de pleistocene valleirand komt frequent veensubstraat voor onder de klei. De Kalkense Meersen zijn een vrij nat gebied maar sinds de installatie van het pompgemaal in 1975 treedt er toch elk jaar een duidelijke droge periode op in de zomer.
Het gebied werd in 1996 door de Vlaamse overheid volledig erkend als natuurgebied. De Kalkense Meersen maken deel uit van het EG-habitatrichtlijngebied ‘Schelde- en Durmeëstuarium van de Nederlandse grens tot Gent’ en het maakt bijna volledig deel uit van het vogelrichtlijngebied ‘Durme en de Middenloop van de Schelde’. De officiële gebieden van het VEN (Vlaams Ecologisch Netwerk) 1e fase behoren tot ‘De Vallei van de Boven Zeeschelde van Kalkense Meersen tot Sint-Onolfspolder’. De GEN (Grote Eenheden Natuur) en GENO (Grote Eenheden Natuur in Ontwikkeling) gebieden zijn respectievelijk 111 ha en 166,5 ha, wat in totaal een VEN-oppervlakte van 277,5 ha betekent (Degezelle et al. 2004). Op het gewestplan staat 111 ha ingekleurd als natuurreservaat, 486 ha als agrarisch gebied met ecologisch belang en 223 ha als valleigebied (Degezelle et al. 2004). Ondertussen is begin 2010 een GRUP (Gewestelijk ruimtelijk uitvoeringsplan) goedgekeurd dat het hele gebied zal inkleuren als natuurgebied en VEN op het gewestplan.
Het studiegebied van dit onderzoek beperkt zich binnen de Kalkense Meersen tot Broekmeers, Molenmeers, Scherenmeersen, Langendonk en Belham (fig. 3.1). In deze gebieden broeden de meeste weidevogels.
Broekmeers is één van de grootste open graslandcomplexen van de Kalkense Meersen (100 ha) en tevens één van de intensiefst bewerkte delen uit het gebied. Aangezien het merendeel van de percelen in eigendom is van landbouwers met melkkoeien of runderen worden ze nog sterk bemest. Als gevolg bestaat de vegetatie er vooral uit grassen en is ze vrij soortenarm. Broekmeers is het deelgebied met hoofdzakelijk bemeste percelen met beheersovereenkomsten. (23 ha beheersovereenkomsten in 2009). Molenmeers, Scherenmeersen, Langendonk en Belham worden verder in de tekst samen genoemd als de reservaatzone (330 ha). Deze deelgebieden bevatten immers een groot deel van de percelen die beheerd worden door Natuurpunt (115 ha). Deze percelen staan ingekleurd als natuurgebied en VEN en daar geldt een mestbeperking. In de reservaatzone zijn ook beheersovereenkomst afgesloten voor weidevogels (35 ha in 2009), maar deze percelen liggen meer verspreid over het hele gebied in vergelijking met Broekmeers.
De vegetatie bestaat voor het grootste deel uit vochtige graslanden, zowel soortenarm als soortenrijk (waaronder goed ontwikkelde dotterbloemgraslanden) maar er komen ook verruigde hoekjes, riet, open water, knotbomen, aangeplante populierenbosjes en enkele akkers voor.
De Kalkense Meersen is een deelgebied van het geactualiseerde Sigmaplan en conform het MWeA zal het volledig ingericht worden als ‘wetland’. In het MWeA wordt voor de Kalkense meersen een combinatie van weidevogelgebied (180-200 ha, zilverschoon- en kamgraslanden) en gevarieerd meersengebied (dotter- en vossenstaartgraslanden afgewisseld met kleinere oppervlaktes eutroof water, rietland, grote zeggevegetaties en voedselrijke ruigten) vooropgesteld als gewenste ontwikkeling. Voor specifieke habitatdoelstellingen en figuren wordt verwezen naar Van Ryckegem et al. 2010.
Broekmeers zal de kern van het weidevogelgebied vormen en krijgt een invulling met zilverschoon- en natte kamgraslanden. Molenmeers vormt naast Belham één van de twee kerngebieden voor het ontwikkelen van dottergraslanden. Momenteel zijn daar al goed ontwikkelde dottergraslanden aanwezig en een gericht beheer moet deze in staat stellen zich verder te ontwikkelen en uit te breiden. De visvijvers worden ingericht tot een complex van eutrofe plassen met goed ontwikkelde oeverzones. In Scherenmeers zal een moeraszone ontwikkeld worden waarin grote zegge- en verlandingsvegetaties, rietpartijen en open water elkaar afwisselen. Langendonk zal de overgangszone van het weidevogelgebied naar de dottergraslanden van Belham vormen. De glanshavergraslanden zullen verder ontwikkeld worden waarbij ruimte is voor kleine landschapselementen zoals knotbomen. Aansluitend op de ‘vliegerkesweide’ zal de ‘grutto-weide’ ontwikkeld worden tot plas-dras ten behoeve van de weidevogels (Van Ryckegem et al. 2010).
3.1.2 Het Aubroek
Het Aubroek ligt op de linker Schelde-oever en volledig op het grondgebied van Berlare (fig. 3.2). De afstand tussen de Kalkense Meersen en het Aubroek bedraagt ongeveer 6 km in vogelvlucht. Het gebied is ongeveer 110 ha groot en maakt deel uit van de Scheldebroeken, een laaggelegen cultuurlandschap op alluviale bodem. De vegetatie bestaat er voornamelijk uit soortenarme graslanden, enkele akkers en populierenaanplanten. Landbouwactiviteiten zijn hier nog iets intensiever dan op Broekmeers omdat er meer melkveehouders actief zijn. Er zijn slechts enkele reservaatperceeltjes in beheer bij vzw Durme (iets minder dan 4 ha). Het Aubroek is het gebied met hoofdzakelijk bemeste percelen zonder beheersovereenkomsten en dient als referentiegebied van Broekmeers om het bruto territoriaal succes van grutto’s te vergelijken.
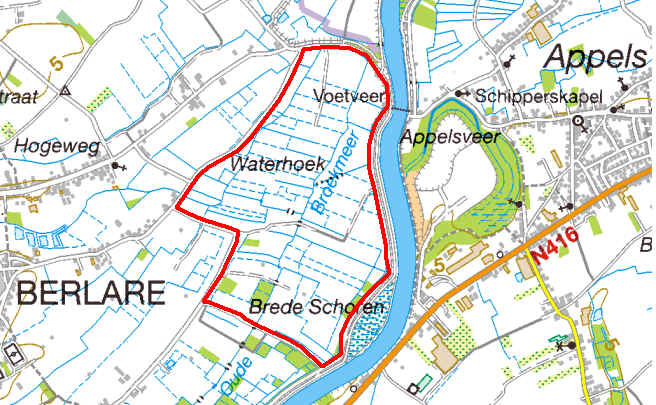
Figuur 3.2. Het Aubroek
3.2 Onderzoeksoorten (gebaseerd op Svensson et al. 1999, Beintema et al. 1995)
3.2.1 Grutto (Limosa limosa)
Grutto’s zijn grote, slanke steltlopers die een lengte kunnen bereiken van 40 tot 44 cm (inclusief hun 7,5 tot 12 cm lange snavel). Vrouwtjes zijn gemiddeld 5% zwaarder en hebben snavels die gemiddeld 15% langer zijn dan die van mannetjes. Volwassen mannetjes zijn in de zomer roestrood op de kop, nek, borst en het voorste deel van de buik en flanken. De overige delen van de onderzijde zijn wit. Op de kruin hebben ze overlangse, donkere streepjes en de delen van de borst en de buik zijn dwars gebandeerd. De snavel heeft een oranjeroze basis met een zwarte punt en de poten zijn zwart. De vrouwtjes zijn over het algemeen iets valer en lichter van kleur maar in alle kleden hebben zowel mannetjes als vrouwtjes een opvallend vliegbeeld met een brede witte vleugelstreep en een zwarte eindband op de witte staart. In de winter hebben grutto’s een bleek grijsbruine bovenzijde en een witte onderzijde.
In de broedtijd zijn grutto’s zeer luidruchtige vogels en ware luchtacrobaten. De baltsvlucht bevat ondermeer het kantelen of wiekelen, waarbij een grutto zijn vleugels met verschillende frequentie beweegt en zich schoksgewijs van de ene op de andere zijde gooit. Tijdens het kantelen roepen ze het melodieuze twee- of driedelige ‘grutto, grutto’. Daarna laten ze zich als een pijl recht naar beneden vallen om na de landing hun vleugels met helderwitte onderzijde enkele seconden omhoog te houden. Dit laatste wijst erop dat het mannetje een territorium heeft.
Grutto’s verkiezen vochtige gebieden en het broedhabitat omvat hoogveen, natte heide, vochtige hooilanden, extensief begraasde weilanden, moerassen en gemaaide rietlanden. Als nestplaats verkiezen de grutto’s plekken met wat langer gras. Mannetjes maken verschillende nestkuiltjes waarvan het vrouwtje er dan één uitkiest. De toppen van het gras worden om het nest gebogen zodat er vaak een soort gat in de vegetatie zit. In dit nestkuiltje verschijnen vanaf eind maart de eerste legsels die bestaan uit vier olijfgroene of bruine eieren met onduidelijke vlekken. De eieren worden ongeveer 24 dagen bebroed. Enkel bij vroegtijdige verstoring wordt een vervolglegsel gelegd. Na 30 tot 35 dagen zijn de jongen vliegvlug. Eieren en jongen worden heftig verdedigd en gruttoparen werken dikwijls samen om predators weg te jagen. Bij verstoring van de jongen laten ze dikwijls hese, langgerekte alarmroepen horen.
Met hun tere snavel foerageren grutto’s liefst in zachte bodem. Met het beweegbare topje van hun bovensnavel zijn ze in staat om prooien uit de grond te grijpen. In het broedseizoen leven volwassen grutto’s vooral van regenwormen, insecten en hun larven (emelten). De jongen zijn nestvlieders en eten vooral insecten die ze in hoog gras vinden. Dit zijn vooral vliegen en muggen.
Na de broedtijd verzamelen grutto’s zich meestal in grotere groepen om vanaf juli ons land te verlaten. De meeste vogels trekken via Spanje, Portugal en Marokko naar de overwinteringsgebieden in Afrika (kusten en rivieren van Senegal, Gambia, Mali,...). De eerste grutto’s arriveren terug in België tegen eind februari.
3.2.2 Kievit (Vanellus vanellus)
Aangezien de kievit ook geïnventariseerd is wordt hier een beknopt overzicht gegeven van de kenmerken van deze soort. Kieviten zijn 28 tot 31 cm lang (2,5 cm lange snavel inclusief). Ze hebben een opvallende kuif en het verenkleed heeft een zwart-witpatroon waardoor ze gemakkelijk te herkennen zijn. Mannetje en vrouwtje lijken sterk op elkaar maar zijn te onderscheiden aan de kortere kuif, de valer gekleurde bovenzijde en de spitsere vleugels van het vrouwtje die bovendien meer wit hebben aan de uiteinden. In alle kleden hebben kieviten een zwarte snavel en vleeskleurig tot rode poten.
Mannetjes hebben een opmerkelijke zangvlucht met diverse bewegingen. Deze vlucht omvat o.a. een traject waarbij het mannetje zich steeds na vier à vijf vleugelslagen van de ene op de andere zijde gooit, waardoor de opvallende wit-zwarte onderzijde zichtbaar wordt. Tegelijkertijd brengen de handpennen een ritmisch, zoevend geluid voort.
Kieviten leven in uiteenlopende open gebieden met geen of weinig en lage vegetatie. Ze broeden in moerassen, hoogveengebieden, oeverzones, duinweiden, weilanden, hooilanden en akkers. In de Kalkense Meersen vertaalt dit zich in de wei- en hooilanden en enkele koppels op akkers. Kieviten prefereren kort gras om hun nest in te bouwen. Het mannetje krabt in zijn territorium verschillende nestkuiltjes uit, waarvan het vrouwtje er één kiest. Daar legt ze vanaf half maart in vijf dagen vier olijfbruine, groenige, zwartgevlekte eieren. De broedtijd bedraagt bijna vier weken en nadien zijn de jongen na 35 tot 40 dagen vliegvlug. Bij verlies van legsels of jongen kunnen kievitsvrouwtjes tot vier vervolglegsels produceren.
Het voedsel van kieviten bestaat voornamelijk uit op de grond levende insecten en hun larven, regenwormen, slakjes en een beetje plantaardig materiaal. De jongen zijn ook nestvlieders, zoals de gruttojongen, maar ze zijn wat trager en moeten het vooral hebben van langzamere prooien die zich op de grond bevinden zoals kevers.
Vanaf juni trekken kieviten weg naar het Iberisch schiereiland of Noord-Afrika om te overwinteren. Bij zachte winters blijft een deel in ons land.
3.3 Het weer in 2009
Het weer kan op verschillende wijzen een invloed hebben op het broedsucces van weidevogels: op de legdatum, het uitkomstsucces van legsels en vooral op de groei en overleving van kuikens. Daarnaast heeft het weer ook invloed op het verloop van agrarische werkzaamheden zoals het tijdstip waarop gemaaid wordt. Daarom wordt hieronder een korte beschrijving gegeven van het weer tijdens het broedseizoen 2009, aan de hand van de klimatologische maandoverzichten van het KMI.
Maart 2009 werd gekenmerkt door normale waarden van het neerslagtotaal, de zonneschijnduur, de gemiddelde windsnelheid en temperaturen. April was uitzonderlijk warm. De gemiddelde temperatuur bedroeg 12,5 °C i.p.v. 9 °C normaal. Ook de zonneschijnduur was voortreffelijk: 297 uur t.o.v. 178 uur normaal. Het neerslagtotaal was normaal maar er werden wel 10 onweersdagen vastgesteld tegenover 6.6 normaal. Van de 10e tot de 16e was er elke dag sprake van onweer. Ook mei werd gekenmerkt door een abnormaal hoge gemiddelde temperatuur: 14,4 °C tegenover 12,7°C normaal. De zonneschijnduur lag iets lager dan normaal en de gemiddelde neerslag was normaal. In juni heersten normale waarden van temperatuur, neerslag en zonneschijnduur.
Als het klimatologisch overzicht van de lente 2009 (maart t.e.m. mei) bekeken wordt is duidelijk dat dit een uitzonderlijk warm seizoen was (tweede plaats in de rij van de warmste lentes sinds de metingen). De gemiddelde temperatuur bedroeg 11,2 °C tegenover 9,5 °C normaal. Met 158 l/m2 t.o.v. 196 l/m2 was de lente ook vrij droog. De waarden van zonneschijnduur waren normaal.
3.4 Broedparen en bruto territoriaal succes
Tijdens het broedseizoen 2009 (half maart – midden juni) werd in elk deelgebied minstens één maal per week naar broedparen van kieviten en grutto’s gezocht. Om verstoring zoveel mogelijk te vermijden werd geopteerd om de volledige inventarisatie uit te voeren vanaf de bestaande paden en wegen die de gebieden doorkruisen. Nesten bezoeken verhoogt immers de kans op predatie doordat geur of zichtbare sporen achterblijven (Teunissen et al. 2005). Aangezien het niet evident is om op deze manier nesten visueel te ontdekken is vooral gelet op het gedrag van de vogels. Kieviten en grutto’s zijn tijdens het broedseizoen zeer luidruchtige vogels en aan de manier van lopen, vliegen, landen en alarmeren is dikwijls op te maken in welk broedstadium ze zich bevinden. Voor meer details wordt verwezen naar de Ecologische Atlas van de Nederlandse Weidevogels (Beintema et al. 1995) en Instructie Alarmtellingen (Nijland & van Paassen 2007). Nadien is tijdens de kuikenfase aan de hand van alarmtellingen nagegaan hoeveel van de oorspronkelijke broedparen van grutto nog alarmeerden op het einde van het broedseizoen. Zo krijgt men een schatting van het aantal broedparen dat succesvol één of meerdere jongen heeft groot gebracht. Dit wordt het bruto territoriaal succes (BTS) genoemd en het geeft een index voor het broedsucces. Het levert echter geen volledige meting op van het broedsucces, omdat hiervoor een aanname nodig is over het aantal levende kuikens per alarmerend paar en de overleving daarvan tussen de alarmtelling en de vliegvlugge leeftijd (Nijland 2002). De gevolgde methode is volledig analoog aan de methode die beschreven wordt door Nijland & van Paassen (2007), behalve dat het veld niet doorkruist werd volgens een vaste looproute maar dat op de paden is gebleven. Om deze reden was het niet altijd mogelijk een onderscheid te maken tussen koppels met nest en koppels met jongen. Als dat het geval was werden deze aantallen geschat. Het BTS wordt berekend door het aantal koppels met jongen in de fladderweken (week 21: 20-27 mei) te delen door het aantal territoria en vervolgens deze waarde te vermenigvuldigen met honderd. Met de fladderweken wordt de periode bedoeld waarin de meeste kuikens bijna vliegvlug zijn. Voor het aantal territoria moet het op één na hoogste aantal getelde territoria gebruikt worden.
Deze methode is echter niet goed toepasbaar op de kievit aangezien deze zijn kuikens dikwijls alleen laat in de laatste periode voor het vliegvlug worden en dat het broedseizoen bij deze soort erg langgerekt is. Voor de kievit is daarom enkel het aantal broedparen bepaald.
Tijdens de inventarisatie is gebruik gemaakt van een verrekijker (Kite Petrel) en een telescoop (Swarovski Habicht AT80). De gegevens werden genoteerd op kleurenorthofoto’s die gedownload werden van de website www.gisoost.be.
3.5 Aandeel niet gemaaid grasland
Gedurende het broedseizoen 2009 werden de maaidata bijgehouden voor een deelgebied van Broekmeers en een deelgebied van het Aubroek. Dit gebeurde tijdens de inventarisaties van de grutto- en kievitkoppels. Aangezien het merendeel van de percelen gemaaid werd in de periode tussen twee inventarisaties, is de datum genoteerd wanneer geconstateerd werd dat bepaalde percelen gemaaid waren. Er werd geen rekening gehouden met hergroei (percelen waar na maaien het gras terug hoger staat dan 15 cm) aangezien dit ook minder geschikt is qua voedselvoorziening en beschutting dan ongemaaide percelen (Schekkerman 1997). De deelgebieden omvatten de belangrijkste percelen waarop alle grutto’s en de meeste andere weidevogels broedden. Het deelgebied in Broekmeers was 52,65 ha groot en dat in het Aubroek 50,27 ha. De oppervlakte van de individuele percelen werd afgeleid uit het kadasterplan dat beschikbaar is op www.gisoost.be. Nadien werd voor de verschillende maaidata berekend hoeveel procent van de oorspronkelijke oppervlakte grasland nog niet gemaaid was. Dit nog niet gemaaid grasland wordt ook ‘kuikenland’ genoemd.
3.6 Historisch verloop gruttopopulatie
Om een zo compleet mogelijk historisch verloop van de gruttopopulatie op te stellen uit de regio van de Kalkense Meersen is gezocht naar alle mogelijke broedgegevens van grutto’s in die regio. Er werd zowel gezocht naar het jaarlijks aantal broedparen als naar de broedlocaties. Een deel van de informatie is afkomstig van gegevens die andere personen reeds verzameld hadden (waaronder Peter Claus, Rudi Clinckspoor, Rudi Van Onderbergen en Gunter De Smet), de rest uit broedvogelinventarisaties, artikels uit lokale tijdschriften (Swaeneblomme), reconstructies van losse waarnemingen, mondelinge mededelingen, enzovoort. Met dit lange termijn overzicht kunnen trends in populatiegrootte en eventuele veranderingen in de broedlocaties aangetoond worden. Het oorspronkelijke opzet was om hetzelfde te doen voor de kievit maar dit was niet mogelijk aangezien er niet genoeg gegevens beschikbaar waren.
3.7 Voedselaanbod onder de vorm van regenwormen
Op 15 en 22 april 2010 werd het voedselaanbod voor volwassen grutto’s onder de vorm van regenwormen onderzocht in Broekmeers en Molenmeers. Er werden vijf stalen genomen in Broekmeers en vijf in Molenmeers. De stalen van Broekmeers werden genomen op percelen die elk jaar tweemaal bemest worden met drijfmest. De stalen in Molenmeers werden genomen op Natuurpuntpercelen waarop al minstens 6 jaar een verschralingsbeheer wordt uitgevoerd met nulbemesting. De percelen werden zo gekozen dat bodem en hydrologie gelijkaardig waren om andere effecten dan de bemesting uit te sluiten. De bodemtextuur van alle percelen bestond uit zware klei en de hydrologie ging van nat op Broekmeers tot zeer nat in Molenmeers. Methoden voor het verzamelen van regenwormen uit de bodem zijn van diverse aard, gaande van chemische methoden met formaldehyde, AITC, mosterdpoeder, detergenten, elektrische methoden (octet), hittebehandelingen (Kempson) tot het uitgraven en sorteren van de bodem met de hand (Čoja et al. 2008).
De methode die toegepast werd in dit onderzoek is een combinatie van een chemische methode met allyl isothiocyanate (AITC) en het handmatig uitsorteren van de bodem. Een combinatie van een chemische methode met het uitsorteren leidt tot de beste schattingen van de regenwormenpopulatie (Pelosi et al. 2009). AITC is een natuurlijk afbraakproduct van glucosinolaten dat dikwijls gevonden wordt in kruisbloemige groenten en verantwoordelijk is voor de scherpe smaak ervan. Het komt bijvoorbeeld voor in kool, mosterd en bieten (Zhang & Thalalay 1994). AITC irriteert de regenwormen waardoor ze naar de oppervlakte komen. Er werd gekozen voor AITC omdat deze stof minder schadelijke gevolgen heeft voor de bodem en niet-doelsoorten dan formaldehyde. AITC breekt snel af in de bodem, het heeft een halveringstijd van 80 tot 120 uur (Borek et al. 1995). De methode is gemakkelijk te standaardiseren in tegenstelling tot de mosterdmethode (Pelosi et al. 2009) en ze is bovendien gemakkelijk toepasbaar in het veld.
Alvorens te vertrekken naar de onderzoeksplaats werden in het labo (Universiteit Gent) tien stockoplossingen gemaakt, één voor elk proefvlak. Daarvoor werd 2 ml AITC (96%) toegevoegd aan 16 ml isopropanol (99,9%). Isopropanol werd gebruikt als emulgator omdat AITC niet oplost in water. Aangezien AITC gevoelig is voor licht werden de stockoplossingen getransporteerd in donkere potjes. In het veld werd de stockoplossing toegevoegd aan 20 l water om zo een 1mM oplossing te bekomen. Deze concentratie wordt als optimaal beschouwd (Čoja et al. 2008). Vervolgens werd een houten kader van 50 x 50 cm op de bodem gelegd waarbinnen het grootste deel van de vegetatie verwijderd werd zodat de kale bodem zichtbaar was. De 20 l AITC oplossing werd, na goed schudden, met een gieter in twee keer op het proefvlak gegoten. Nadat de eerste 10 l oplossing in de bodem was gedrongen werden gedurende tien minuten alle wormen verzameld die aan de oppervlakte kwamen. Vervolgens werd de laatste 10 liter oplossing op het proefvlak gegoten om opnieuw gedurende tien minuten wormen te verzamelen. De ingezamelde wormen werden bewaard in ethanol (99,9%). Van in twee gebroken wormen werd maar één helft ingezameld. Zeer kleine juveniele wormpjes (nog wit van kleur) werden niet gecollecteerd. Na deze chemische methode werd met een spade een stuk bodem uitgegraven van 25 x 25 cm en een diepte van 15 cm. Alle resterende wormen die zich in deze blok grond bevonden werden verzameld. Als laatste werd een staal van de bodem meegenomen om in het labo de pH te meten. De hele procedure nam per proefvlak ongeveer 45 minuten in beslag.
Terug in het labo werden alle wormen geteld en gewogen. Aangezien de wormen onmiddellijk na het veldwerk gewogen werden representeert het gewicht de eigenlijke biomassa van de wormen plus de maaginhoud. Verder in de tekst zal de term biomassa gebruikt worden voor deze som. De weegschaal (Kern EMB 600-2) was tot op 0,01 g nauwkeurig en de massa van wormen die minder wogen dan 0,01 g werd gelijk gesteld aan 0,01 g. De massa van een in twee gebroken worm werd vermenigvuldigd met factor 2. Nadien werden alle waarden omgezet naar m². Voor het testen van een verschil in het totale aantal wormen tussen de bemeste en onbemeste proefvlakken werd een veralgemeend lineair model gebruikt. Het verschil in totale biomassa en pH werd geanalyseerd met een algemeen lineair model. Deze veralgemeende en algemene lineaire modellen werden uitgevoerd in het statistische programma SAS ® (versie 9.2). Om de gemiddelde massa van de wormen te vergelijken werd een permutationele ANOVA uitgevoerd met bemestingsgraad als fixed factor en proefvlak genest binnen bemestingsgraad als random factor, gebaseerd op Euclidische afstanden en 99.999 permutaties van residuelen onder een gereduceerd model. Deze test werd uitgevoerd in Primer (Anderson et al. 2008).
3.8 Vegetatiehoogte
Op 30 maart en 3 april 2010 werd respectievelijk in Broekmeers en Molenmeers de hoogte van de vegetatie opgemeten. Deze data stemmen overeen met de periode waarin grutto’s hun nestplaats kiezen en beginnen aan de bouw van een nest. In Broekmeers werden de metingen uitgevoerd op 30 plaatsen, willekeurig verspreid over percelen die bemest worden. In Molenmeers werd de vegetatie opgemeten op 30 willekeurige plaatsen die in beheer zijn bij Natuurpunt en een verschralingsbeheer kennen. Voor de meting werd een plastieken mapje (31 x 22,5 cm;
22 g) op de vegetatie gelegd. De hoogte van de vegetatie werd vervolgens bepaald door de hoogte, vanaf de bodem gemeten, waarop het plastieken mapje bleef rusten op de vegetatie. Om het verschil tussen de twee groepen te testen werd een algemeen lineair model (Welch’s ANOVA) toegepast via het statistische programma SAS ® (versie 9.2).
3.9 Kruidenrijkdom
Op 27 mei en 16 juni 2009 werd de soortenrijkdom bepaald in verschillende bemeste en onbemeste percelen. De bemeste percelen bevonden zich allemaal in broekmeers, de onbemeste percelen (eigendom van Natuurpunt) bevonden zich op Molenmeers, Langendonk en Belham. De bemeste percelen in Broekmeers worden bijna allemaal minstens twee maal per jaar bemest met drijfmest of kunstmest. De hooilanden die in beheer zijn bij Natuurpunt krijgen een verschralingsbeheer waarbij één- of tweemaal gemaaid wordt. Er werden 19 bemeste proefvlakken geïnventariseerd en 14 onbemeste. Proefvlakken werden willekeurig toegewezen door een kader van 1 x 1 m in het perceel te leggen. Om randeffecten te vermijden werden de proefvlakken niet te dicht tegen de rand van de percelen gelegd. Vervolgens werden alle aanwezige plantensoorten in het proefvlak gedetermineerd. Om het aantal soorten tussen bemeste en onbemeste proefvlakken te vergelijken werd een veralgemeend lineair model gebruikt dat uitgevoerd werd in SAS ® (versie 9.2).

Gruttonest met vier olijfgroene, gevlekte eieren. (Peter Claus)
4 Resultaten
4.1 Broedparen en bruto territoriaal succes
4.1.1 Grutto
De eerste grutto’s arriveerden in de Kalkense Meersen omstreeks 25 februari 2009 (www.waarnemingen.be), maar dit kunnen ook doortrekkers geweest zijn. In tabel 9.1 (zie bijlage I) is het aantal broedparen weergegeven die op de aangegeven data werden waargenomen in de verschillende deelgebieden. De eerste territoria werden vastgesteld op 9 april 2009. Toen werden er in de reservaatzone, met uitzondering van Molenmeers, 6 territoria opgetekend en in Broekmeers 4. Het maximum aantal territoria dat waargenomen werd bedroeg 14, zowel voor de reservaatzone als voor Broekmeers. De laatste grutto’s werden gezien op 16 juni. Dit waren 4 alarmerende koppels in Broekmeers.
Het totaal aantal broedparen in de Kalkense Meersen (in 2009) wordt op 24 geraamd: op 22 april werden 21 broedparen opgetekend en er bevonden zich nog 3 broedparen buiten het onderzoeksgebied van de studie (2 Oude Schelde, 1 Wijmeers). Van deze 21 broedparen hadden er 14 hun nest in de reservaatzone en 7 in Broekmeers (situatie voor de verschuiving naar Broekmeers). Dit komt neer op een dichtheid van 6 broedparen (bp) per km² (100 ha) in de reservaatzone en 7 bp/km² in Broekmeers. Een kaartje met de locaties van de nesten is te vinden in bijlage I (fig. 9.1). Vermeldenswaardig is ook een broedkoppel in de Molenveldkouter (net buiten de Kalkense Meersen) dat broedde op een raaigrasperceel, gelegen in een zandig akkergebied.
Een opmerkelijk feit is dat in de week vóór 30 april alle broedparen van de gruttoweide verdwenen zijn! In Belham werden de laatste grutto’s gezien op 15 mei, waaronder één jong dat nog lang niet vliegvlug was. Nadien werden ook geen koppels meer aangetroffen in de ruime omgeving wat doet vermoeden dat die koppels weinig of geen jongen zullen grootgebracht hebben. Daar tegenover staat dat het aantal broedparen in Broekmeers bleef stijgen tot 1 mei met nog een uitschieter op 15 mei. Waarschijnlijk zijn een viertal koppels, afkomstig van de gruttoweide, opnieuw beginnen broeden in Broekmeers. Om die reden werd de berekening van het bruto territoriaal succes (BTS) bemoeilijkt, maar er is geprobeerd hiermee rekening te houden.
Voor Broekmeers bedroeg het aantal koppels in de fladderweken 10 en het op één na hoogste getelde aantal territoria 11. Met deze cijfers bekomen we een BTS van 90,9%. Ik veronderstel echter dat er op 20 mei nog 4 koppels aanwezig waren met een nest omdat ze na hun poging op de gruttoweide opnieuw begonnen zijn in Broekmeers. Als we dit in rekening brengen dan bedraagt het aantal koppels met jongen in de fladderweken 6. Om ook geen territoria mee te rekenen van koppels afkomstig uit de reservaatzone moeten we het op één na hoogste aantal territoria nemen van vóór 30 april. Het aantal territoria bedraagt dan 7. Als we deze waarden gebruiken bedraagt het BTS 85,7%.
Als we, zoals hierboven vermeld, veronderstellen dat er zich een viertal koppels uit de reservaatzone opnieuw gevestigd hebben in Broekmeers rond 30 april, dan kunnen we er zo goed als zeker vanuit gaan dat het deze 4 koppels zijn die nog alarmeerden op 16 juni. Alle andere koppels zijn dan normaal al vertrokken naar nattere gebieden omdat de Kalkense meersen dan te droog worden (pers. med. Peter Claus). 16 juni valt voor deze vier koppels ongeveer in de fladderweken zodat het BTS van deze koppels 100% bedraagt. Voegen we dit samen met de oorspronkelijke koppels van Broekmeers dan komen we aan 11 territoria waarvan er nog 10 alarmeerden in de fladderweken. Dit resulteert opnieuw in de BTS-waarde van 90,9%.
In de reservaatzone bedraagt het BTS 21,4%. Als aantal alarmerende koppels met jongen in de fladderweken is 3 genomen aangezien op 20 mei slechts 3 van de 4 koppels in Molenmeers jongen had. Het aantal territoria bedroeg 14, dit is het hoogste aantal getelde territoria. Merk op dat het op één na hoogste aantal getelde territoria, 8 broedparen op 24 april (zie bijlage I, tabel 9.1), niet representatief is aangezien er toen niet geteld werd in Langendonk.
In het Aubroek werden de eerste territoria waargenomen op 28 maart, het laatste koppel werd gezien op 13 juni (zie bijlage I, tabel 9.2). Het aantal broedparen wordt geschat op 5. Dit betekent een dichtheid van 5 bp/km². Met twee koppels op 23 mei en 4 territoria bedraagt het BTS 50%.
4.1.2 Kievit
Op 9 en 10 april 2009 werden 30 koppels kievit geteld in de reservaatzone en 18 koppels in Broekmeers. Dit zijn echter enkel de koppels die in grasland broedden. Er broedden ook enkele koppels op de weinige akkers in het gebied waardoor we kunnen spreken van minstens 55 broedkoppels kievit in het studiegebied. Dit levert een dichtheid van 13 bp/km² op. Net zoals alle gruttokoppels, verdwenen ook alle aanwezige kievitparen van de gruttoweide in de week vóór 30 april.
In het Aubroek werden maximaal 18 broedende koppels kievit waargenomen waarvan minstens een vijftal op akkers. De dichtheid bedraagt 16 bp/km².
4.2 Aandeel niet gemaaid grasland
Figuur 4.1 geeft een verloop weer van het aandeel niet gemaaid grasland in Broekmeers en het Aubroek. De data weerspiegelen niet altijd de exacte datum van de maaiwerkzaamheden aangezien sommige percelen een paar dagen voor de inventarisatie gemaaid werden. De data tonen dus eerder op welke dag geconstateerd werd dat bepaalde percelen gemaaid waren.
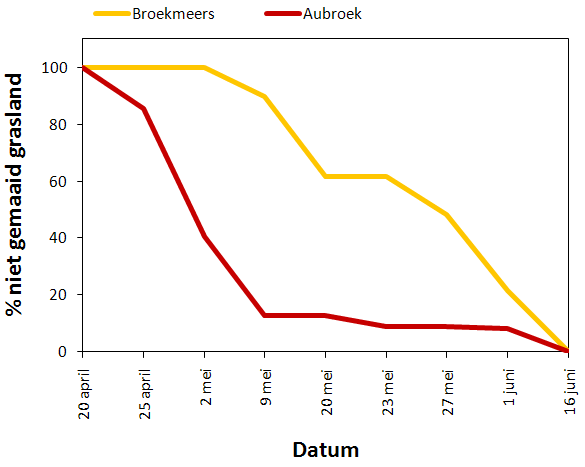
Figuur 4.1. Verloop van het aandeel niet gemaaid grasland voor een deelgebied in Broekmeers (geel) en het Aubroek (rood).
In het Aubroek werden de eerste percelen gemaaid in de week van 25 april 2009. Toen werd ongeveer 14% van het grasland gemaaid. Tijdens de twee weken die volgden werd het merendeel van de andere percelen ook gemaaid waardoor op 9 mei slechts 12,7% niet gemaaid grasland overbleef. Tussen 20 mei en 1 juni werd nog 4% gemaaid en na 16 juni was alles gemaaid.
In Broekmeers werd pas voor het eerst gemaaid in de week van 9 mei. Er werd toen ongeveer 10% van de oppervlakte gemaaid. Tussen 9 en 20 mei werd nog een deel gemaaid met als resultaat nog 61,6% niet gemaaid grasland. In de week van 27 mei werd 13,6% gemaaid en in de periode daaropvolgend nog eens 26,4% waardoor op 1 juni nog 21,5% niet gemaaid grasland overbleef. Op 16 juni werden alle resterende percelen gemaaid.
4.3 Historisch verloop gruttopopulatie
4.3.1 Aantal
De oudste indicaties over het voorkomen van grutto’s in de Kalkense Meersen zijn afkomstig uit het boek ‘Het kasteel en de meersen’ (van Damme & Lostrie 1987). Hierin wordt vermeld dat grutto’s vroeger zo abundant waren dat hun eieren verzameld en verkocht werden voor de tafels der rijken. De eerste bekende broedgegevens dateren uit 1967. Toen werden twee broedgevallen vastgesteld in Wijmeers. In de daaropvolgende jaren werden telkens 2 à 3 koppels waargenomen met een maximum van 4 in 1975 (fig. 4.2). In de periode 1982 - 1984 waren elk jaar 4 broedende koppels aanwezig. Concrete gegevens van de jaren 1970, 1971, 1977, 1978 en 1986 ontbreken. 1987 liet 6 broedparen optekenen en vanaf dan steeg de gruttopopulatie gestaag tot een maximum van 30 broedparen in 2002. Twee knikken in de stijgende lijn zijn te vinden in 1993 en 1997 met respectievelijk 13 en 18 broedparen. De laatste jaren (2003 tot 2008) stagneert de populatie rond 25 koppels. In 2009 werden 24 koppels geteld.
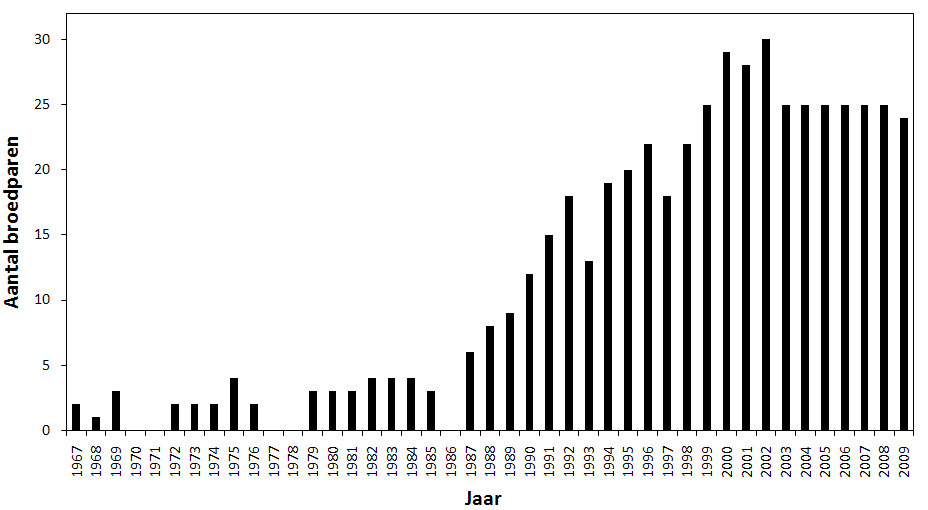
Figuur. 4.2. Aantal broedparen in de Kalkense Meersen (1967 – 2009). Jaren waarvoor geen balk aanwezig is wijzen op ontbrekende gegevens.
Ondanks de stagnatie in de Kalkense Meersen zelf, is er de laatste tijd een uitbreiding van de gruttopopulatie te merken naar gebieden in de buurt (hierna: ‘randgebieden’) zoals de Scheldebroeken in Berlare (Aubroek, Paardenweide) en de weiden rond het station van Schellebelle. In het Aubroek werden vanaf 2001 2 broedende koppels waargenomen en dit aantal steeg de laatste jaren tot 5 koppels (pers. med. R. van Onderbergen). Ook op de Paardenweide werd het eerste koppel waargenomen in 2001. In 2005 liep dit aantal op tot 5 koppels maar in 2009 trof men echter maar 3 koppels meer aan (pers. med. D. De Mesel). In Schellebelle kwamen sinds 1998 maximaal 4 koppels tot broeden. De laatste jaren is dit aantal teruggelopen tot 2 koppels (pers. med. P. Claus). De broedgegevens van de randgebieden worden weergegeven in tabel 9.3 (zie bijlage II).
Als men het verloop van het totale aantal koppels uit deze randgebieden bekijkt, is zelfs een stijging waar te nemen tot en met 2005 (fig. 4.3). In 2005 kwamen bijna dubbel zoveel koppels tot broeden dan in 2002. De knik in 2004 is te wijten aan ontbrekende gegevens van de Paardenweide in 2004. Daarom is het aantal koppels van de Paardenweide als 0 genomen terwijl dit waarschijnlijk niet het geval was.
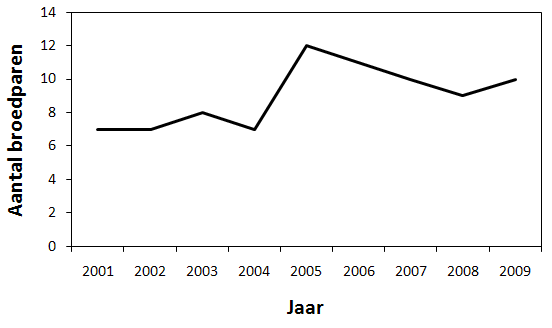
Figuur 4.3. Totaal aantal broedparen in de randgebieden (2001 – 2009).
4.3.2 Locatie
Informatie over de locatie van broedgevallen was minder voorhanden dan de informatie over het aantal broedparen. Figuur 4.4 geeft over verschillende jaren weer hoeveel koppels van de totale populatie in de betreffende deelgebieden broedden. De eerste beschikbare gegevens dateren uit 1967. Toen werden 2 broedgevallen vastgesteld in Wijmeers en tot en met 1975 was er ook enkel sprake van broedgevallen in Wijmeers. In het jaar 1979 kwam daar 1 koppel op Langendonk bij. In 1982 werden 4 koppels waargenomen, 1 op Langendonk, 2 op Molenmeers en 1 op Broekmeers. Sinds 1982 werden tevens geen koppels meer waargenomen in Wijmeers tot recent in 2008 en 2009. Tussen 1985 en 1988 begon de populatie te groeien maar de nieuwe koppels bleven zich enkel vestigen op Langendonk, Molenmeers en Broekmeers. In 1989 werd Scherenmeersen op slag het belangrijkste deelgebied met vijf koppels. Op Langendonk broedde daarentegen geen enkel koppel meer. In de periode 1990 - 1995 kwam Belham erbij en steeg het aantal koppels per deelgebied gelijkmatig waardoor de grutto’s in 1996 met ongeveer evenveel koppels (± 4) vertegenwoordigd waren in elk deelgebied. Na de dip van 1997 begon er een koppel te broeden aan de Oude Schelde en schoot het aantal koppels in Molenmeers spectaculair de hoogte in (10 koppels in 2000). Tijdens de jaren nadien (2000 – 2008) daalde dit aantal echter even spectaculair als het gestegen was en Scherenmeersen verloor zijn aandeel volledig. Broekmeers, Langendonk en Belham gingen daarentegen wel vooruit.
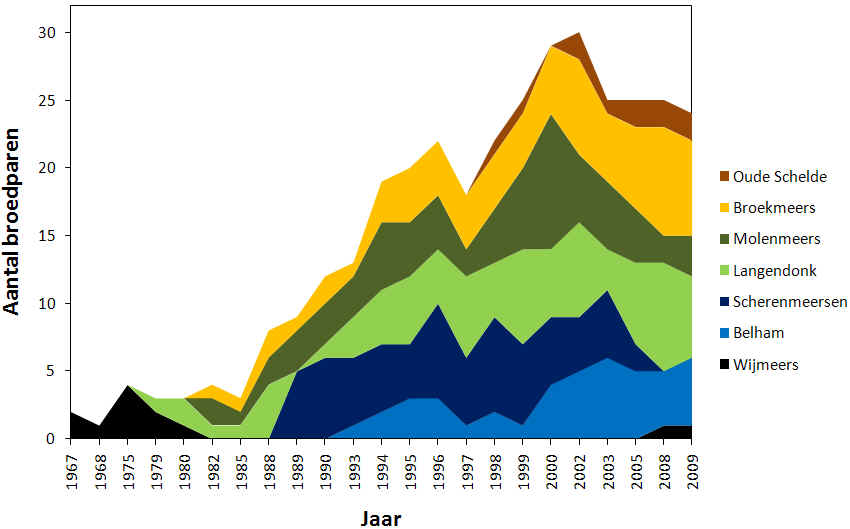
Figuur 4.4. Totaal aantal broedparen in de Kalkense Meersen, opgesplitst per deelgebied (1967-2009).
In figuur 4.5 wordt het aantal koppels van Molenmeers vergeleken met het aantal koppels in Broekmeers. Figuur 4.6 geeft hetzelfde weer maar voor Langendonk + Belham en Scherenmeersen. In 2009 broedden 2 koppels aan de Oude Schelde, 7 koppels in Broekmeers, 3 koppels in Molenmeers, 6 koppels in Langendonk, 5 in Belham en 1 in Wijmeers. De kerngebieden waren dus duidelijk Broekmeers, Langendonk en Belham. Uiteindelijk hebben er op Broekmeers 11 koppels gebroed nadat 4 koppels na hun eerste poging in Langendonk een nieuw nest begonnen zijn in Broekmeers.
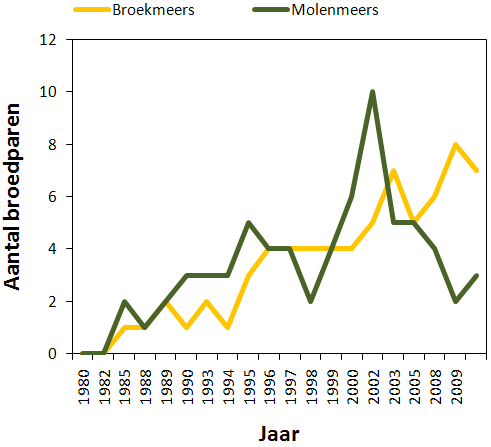
Figuur 4.5. Aantal broedparen in Broekmeers (geel) en Molenmeers (groen) (1980–2009).
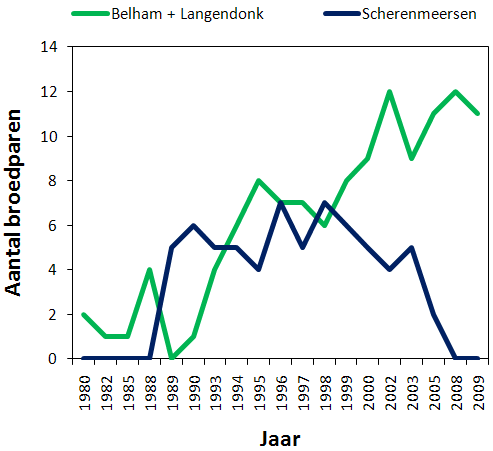
Figuur 4.6. Aantal broedparen in Belham + Langendonk (groen) en Scherenmeersen (blauw) (1980-2009).
4.4 Voedselaanbod onder de vorm van regenwormen
In totaal werden 520 wormen verzameld over alle proefvlakken. Het totaal aantal regenwormen was duidelijk hoger in bemeste proefvlakken (488 m-2 ± 68,22 m-2; gemiddelde ± standaardfout) dan in proefvlakken van niet bemeste percelen (167,2 m-2 ± 27,43 m-2). De aantallen lagen zelfs meer dan dubbel zo hoog (fig. 4.7). Dit verschil was significant (F1,8 = 16,95 ; p = 0,0034).
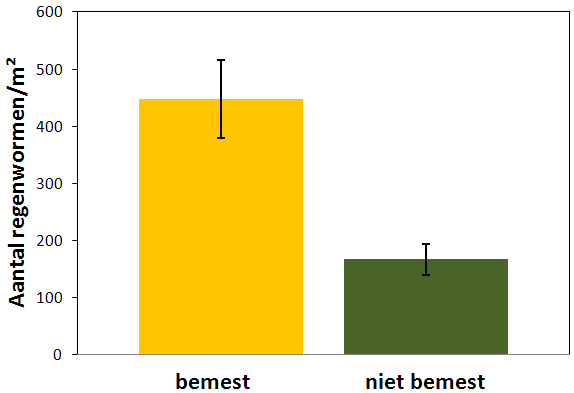
Figuur 4.7. Totaal aantal regenwormen (m-2) in bemeste en niet bemeste proefvlakken (gemiddelde ± standaardfout).
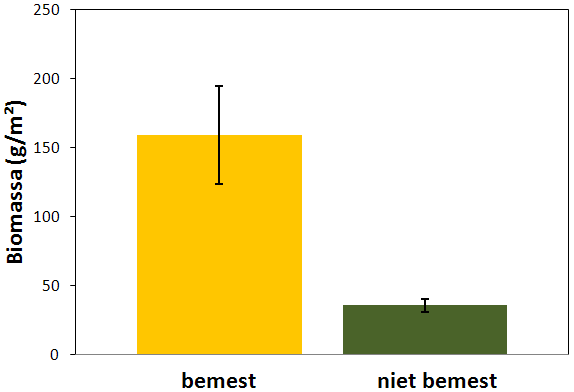
Figuur 4.8. Totale biomassa aan regenwormen (g/m²) in bemeste en niet bemeste proefvlakken (gemiddelde ± standaardfout).
De totale biomassa aan regenwormen was significant hoger in bemeste proefvlakken (F1,8 = 11,9 ; p = 0,0087) en bedroeg daar gemiddeld 159,4 g m-2 (± 35,49 g m-2) tegenover 35,7 g m-2 (± 4,64 g m-2) in niet bemeste proefvlakken (fig. 4.8).
Regenwormen uit bemeste proefvlakken wogen gemiddeld iets meer (0,33 g ± 0,02 g) dan regenwormen uit niet bemeste proefvlakken (0,22 g ± 0,02 g). Dit verschil was niet significant (pseudo-F1;9,68 = 3,21 ; p = 0,13).
De gemiddelde pH was significant lager (F1,8 = 30,55 ; p = 0,0006) in bemeste proefvlakken (6,61 ± 0,07) dan in niet bemeste proefvlakken (7,24 ± 0,09).
4.5 Vegetatiehoogte
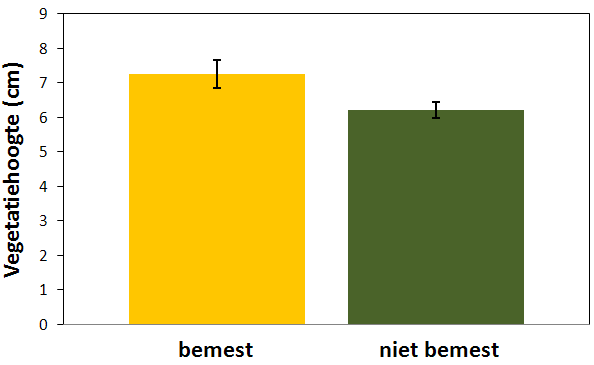
Figuur 4.9. Hoogte (cm) van de vegetatie op bemeste en niet bemeste percelen (gemiddelde ± standaardfout).
De vegetatie was op het moment van de metingen hoger op de bemeste percelen van Broekmeers dan op de onbemeste percelen van Molenmeers. Het verschil tussen de twee groepen was significant (F1;46,5 = 4,95 ; p = 0,031). De gemiddelde hoogte (± standaardfout) van de vegetatie bedroeg in Broekmeers 7,26 cm (± 0,41 cm) en in Molenmeers 6,21 cm (± 0,24 cm) (fig. 4.9).
4.6 Kruidenrijkdom
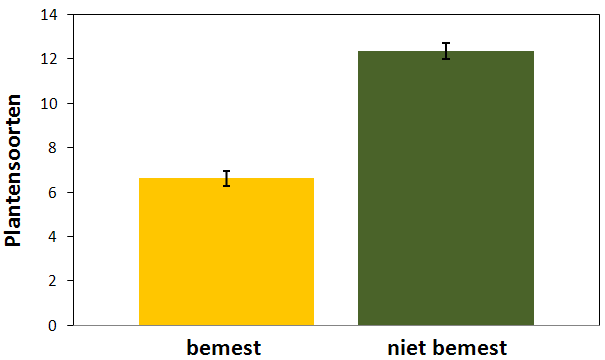
Figuur 4.10. Aantal soorten planten aanwezig in bemeste en onbemeste proefvlakken (gemiddelde ± standaardfout).
In bemeste proefvlakken waren duidelijk minder soorten aanwezig dan in onbemeste proefvlakken (F1,31 = 114,40 ; p < 0,0001). Bemeste proefvlakken bevatten gemiddeld (± standaardfout) 6,63 soorten m-2 (± 0,34 m-2) terwijl in onbemeste proefvlakken bijna dubbel zoveel soorten teruggevonden worden (12,36 m-2 ± 0,37 m-2) (fig. 4.10). De vegetatie in de bemeste proefvlakken wordt vooral gedomineerd door forse grassen. Gestreepte witbol (Holcus lanatus), italiaans raaigras (Lolium multiflorum) en grote vossenstaart (Alopecurus pratensis) worden bijna overal teruggevonden. In onbemeste proefvlakken is de vegetatie veel kruidenrijker. Hier vindt men dikwijls pinksterbloem (Cardamine pratensis), kruipende boterbloem (Ranunculus repens) en echte koekoeksbloem (Silene flos-cuculi). In tegenstelling tot de dichte grasvegetatie op bemeste percelen is er in de meer open vegetatie van onbemeste percelen ook plaats voor enkele zeldzaamheden zoals trosdravik (Bromus racemosus) en kleine ratelaar (Rhinanthus minor) (zie tabel 9.4 in bijlage III voor een lijst met de gedetermineerde soorten).

In de eerste tien dagen na het uitkomen worden gruttojongen nog regelmatig bebroed.
5 Discussie en Conclusies
5.1 Broedparen en bruto territoriaal succes
5.1.1 Grutto
In de Kalkense meersen kwamen dit jaar 24 koppels grutto tot broeden. Dat is één koppel minder dan in de vorige jaren (zie 4.3.1), de populatie blijft dus stabiel. Op het eerste zicht is er nog geen positief effect van het weidevogelproject waar te nemen op de populatiegrootte. Dit is vrij logisch aangezien het weidevogelproject recent werd opgestart in het voorjaar van 2008. Bij een langlevende soort met een beperkte jaarlijkse reproductiecapaciteit zoals de grutto, reageren de aantallen vertraagd op een verbetering van de omstandigheden. Het merendeel van de juveniele grutto’s blijft bovendien tijdens hun tweede kalenderjaar in Afrika en keert pas terug naar hun geboorteplaats als ze twee jaar worden (Groen 1993, Kuijper et al. 2006). Daarenboven moet ook rekening gehouden worden met emigratie, immigratie en de mortaliteit tijdens de trek en in de overwinteringsgebieden. Het is dus onwaarschijnlijk dat op één jaar positieve effecten zichtbaar zijn in het aantalsverloop. Aanwas van jonge vogels die geboren werden in 2008 is pas te verwachten vanaf 2010.
Evaluaties op basis van aantallen zijn dus moeilijk te interpreteren: als het weidevogelproject niet zou leiden tot hogere aantallen wil dit niet noodzakelijk zeggen dat het beheer ineffectief is. De extra geproduceerde kuikens kunnen zich vestigen in andere gebieden. Het omgekeerde is ook mogelijk: dat het project meer vogels naar het gebied lokt, die vervolgens geen broedsucces behalen. Het is zinvoller te kijken of het weidevogelproject iets oplevert voor het broedsucces van de koppels. Hoewel het uiteindelijke doel van het weidevogelproject is om de aantallen te vergroten, is de praktische invulling ervan gebaseerd op het verhogen van het broedsucces. Het broedsucces (of een relatieve maat ervan, zoals het BTS) sluit dus aan bij het veronderstelde mechanisme van de maatregelen en zo kan meer inzicht verkregen worden in de werking van het mechanisme zelf (Schekkerman et al. 2005).
Eerst en vooral moet er op gewezen worden dat de nauwkeurigheid van de schattingen van het BTS beïnvloed kan zijn aangezien de inventarisatie vanaf de paden is gebeurd in plaats van een traject te volgen door de percelen. Er is geprobeerd om dit deels te compenseren via een intensievere inventarisatie dan degene die voorgeschreven wordt in de handleiding van Nijland & van Paassen (2007). Ten tweede is deze handleiding ontwikkeld voor grote weidevogelgebieden in Nederland waar grote aantallen broedkoppels aanwezig zijn. Hierdoor is ze minder bruikbaar bij een klein aantal broedkoppels (pers. med. A. van Paassen). Omwille van bovenstaande redenen moet zeer voorzichtig omgesprongen worden met de interpretatie van de BTS-waarden. Bij het begin van het onderzoek was ik me bewust van deze beperkingen, maar het is de enige manier om op een relatief eenvoudige en goedkope wijze een indicatie te krijgen van het broedsucces van grutto’s. Absoluut kwantificeren van het broedsucces is bij de grutto vrijwel enkel mogelijk door het volgen van individueel herkenbare vogels, toch zeker in de kuikenfase (Schekkerman et al. 2005). Dit kan met behulp van zenders of kleurmerken maar is echter zeer duur en tijdrovend. Een voordeel is wel dat via deze methodes, en in combinatie met de Mayfieldmethode (Mayfield 1975), uitspraken kunnen gedaan worden over de overleving van eieren en jongen apart. Veelal is deze informatie wenselijk aangezien de condities nodig voor de overleving van eieren verschillen met deze voor de kuikenoverleving. Zo kan het beheer hier optimaal op afgestemd worden. Het nadeel van de Mayfieldmethode is wel dat nesten meermaals per week bezocht moeten worden om dagelijkse overlevingskansen van eieren en jongen te kunnen berekenen. Teunissen et al. (2005) vonden dat elk nestbezoek de uitkomstkans gemiddeld met 10% verlaagd. Het werkelijke broedsucces zal op die manier dus ietwat onderschat worden.
In Broekmeers bedroeg het BTS 90,9%, wat wil zeggen dat negen op de tien broedkoppels succesvol één of meerdere jongen hebben voortgebracht. Zo’n hoge waarde is ongezien in Nederland (pers. med. A. van Paassen). Maar het is zeker ook een feit dat de meeste graslanden in Nederland veel intensiever bewerkt worden dan de relatief extensieve graslanden in Broekmeers, te meer door de toepassing van het weidevogelproject. De BTS-waarde van de reservaatzone lag met 21,4% bedroevend laag ondanks het extensieve beheer.
Afgezien van de eventuele onnauwkeurigheid van de BTS-waarden zijn ze toch indicatief voor een duidelijk verschil tussen Broekmeers en de reservaatzone. Verschillende factoren kunnen hierin een rol spelen. Een laag BTS wordt hoofdzakelijk veroorzaakt doordat eieren en/of jongen niet overleven. De voornaamste oorzaken die leiden tot mortaliteit onder grutto-eieren en -jongen zijn landbouwbewerkingen, predatie en slecht weer (Beintema et al. 1995). Aangezien alle kerngebieden van de grutto’s in de reservaatzone onder het weidevogelbeheer of Natuurpuntbeheer vallen (uitgezonderd Belham), werden daar geen landbouwbewerkingen uitgevoerd voor 15 juni. Het weer was bovendien zeer goed in het voorjaar van 2009 (zie 3.3), wat doet vermoeden dat het lage BTS in de reservaatzone veroorzaakt werd door predatie en/of verstoring.
De koppels van de gruttoweide verlieten hun nestplaats vóór eind april en de koppels van Belham rond begin mei.
Als predatie hiervoor verantwoordelijk was, betrof het vooral predatie van de eieren aangezien de uitkomstdatum van gruttolegsels in de Kalkense Meersen meestal rond 1 tot 5 mei ligt (pers. med. P. Claus). In de ecosysteemvisie Kalkense Meersen wordt gewag gemaakt van het voorkomen van verschillende marterachtigen zoals hermelijn (Mustela erminea), wezel (Mustela nivalis), bunzing (Mustela putorius) en steenmarter (Martes foina) (Degezelle et al. 2004), maar ook de vos (Vulpes vulpes) komt er voor. Al deze soorten zijn mogelijke predators van eieren. Ondanks de vaststelling dat zwarte kraaien op grote schaal waarschijnlijk niet de belangrijke predators zijn (Teunissen et al. 2005) zou ik hier wel het belang van kraaien in de predatie/verstoring willen benadrukken. Tijdens het broedseizoen 2009 waren immers steeds een 70-tal zwarte kraaien aanwezig in en rond de gruttoweide. Op de weidepaaltjes die de gruttoweide afbakenen worden wel meer zwarte kraaien gezien maar in 2009 was de groep opvallend groter dan in andere jaren (pers. med. P. Claus). Dat er vlak naast de gruttoweide twee maal per dag gevoederd werd door een lokale burger heeft hier hoogstwaarschijnlijk veel mee te maken. Het voederen had als gevolg dat de kraaien daar steeds aanwezig bleven en dat bovendien meeuwen, die ook als predator bestempeld worden, hierop afkwamen. Bijgevolg werden de grutto’s constant verstoord en werden ze gedwongen voortdurend te alarmeren. Andere vormen van verstoring kunnen afkomstig zijn van de Blokstraat en de Belhamstraat die respectievelijk vlak naast de gruttoweide en het kerngebied op Belham liggen. Op deze wegen passeert nog redelijk veel verkeer van auto’s, maar ook van fietsers en voetgangers. Grutto’s en andere weidevogels zijn storingsgevoelig en volgens Oosterveld & Altenburg (2005) bedraagt de storingsafstand van een tertiaire weg en zelfs van een fietspad ongeveer 100 meter. Het spreekt voor zich dat de grutto’s hier enige hinder van ondervinden.
In het Aubroek, dat nog intensief bewerkt wordt, wordt het aantal broedparen geschat op vijf, waarmee de kleine populatie daar ook al enkele jaren constant blijft. Het BTS bedraagt 50%. Deze waarde is echter onbetrouwbaar en weinigzeggend over het echte broedsucces omdat de steekproef uit zeer weinig koppels bestaat. Door het op één na hoogste aantal territoria te nemen (hier 4 i.p.v. 5 aanwezige territoria) bekomt men direct een hoger BTS dan in werkelijkheid het geval is. De echte BTS-waarde ligt dus vermoedelijk rond 40% of lager. De oorzaak voor dit lage BTS moet in het Aubroek vooral gezocht worden in verliezen door landbouwactiviteiten en predatie. Het grootste deel van het gebied werd reeds zeer vroeg gemaaid (zie 4.2), waardoor vermoedelijk nesten of jongen verloren gegaan zijn. In het Aubroek zijn slechts enkele kleine perceeltjes in beheer bij Vzw Durme en toch broedden hier twee koppels op. Deze percelen worden niet gemaaid voor 15 juni waardoor de nesten gespaard werden. Ondanks hun beperkte omvang en totale oppervlakte zijn ze dus wel degelijk van belang. Een verdere uitbreiding van reservaatpercelen of een inrichting van een weidevogelproject is dus wenselijk.
Vlak naast deze percelen vond ik op 16 mei echter twee gepredeerde grutto-eieren op de weg. Dit wijst erop dat predatie ook een factor is die daar meespeelt, ondanks de openheid van het gebied. In mei en begin juni zijn de beheerde percelen in feite smalle stroken in een zee van gemaaid gras en het is bekend dat grutto’s meer kans hebben om gepredeerd te worden in dergelijke smalle stroken ongemaaid gras (Teunissen et al. 2005). In één van
de eieren was nog een zeer klein embryo aanwezig wat er op wijst dat dit afkomstig was van een zeer laat of vervolglegsel. Op 30 mei vond ik nog een nest met slechts één ei op een reeds gemaaid perceel, wat dus ook een vervolglegsel was. Aangezien er vervolglegsels werden gelegd kan er vanuit gegaan worden dat de vorige nesten verloren gingen door predatie of uitmaaien. Vervolglegsels zijn echter van mindere kwaliteit en hebben een hogere kans om gepredeerd te worden omdat predatie toeneemt naarmate het einde van het broedseizoen nadert (Teunissen et al. 2005). Het is dus zeer belangrijk dat weidevogels zo ongestoord mogelijk hun eerste legsel kunnen uitbroeden om een optimaal broedsucces te behalen.
Om een populatie in stand te houden moeten grutto’s elk jaar gemiddeld 0,5 - 0,8 (waarschijnlijk 0,6 - 0,7) jongen grootbrengen (Schekkerman & Müskens 2000). In termen van BTS betekent dit dat vanaf een percentage van 65% aangenomen wordt dat er voldoende jongen zijn uitgevlogen om sterfte te compenseren (Nijland et al. in voorbereiding.). Op basis van de gegevens kunnen we besluiten dat deze norm enkel gehaald wordt in Broekmeers. Dit is een eerste indicatie dat het weidevogelproject mogelijk een positieve bijdrage levert voor de grutto’s in Broekmeers. Waren alle percelen vroeger gemaaid geweest, dan zouden wellicht minder nesten en kuikens overleefd hebben waardoor niet zo’n hoog BTS zou gemeten worden. In de reservaatzone werd het positief effect van het project en het natuurbeheer teniet gedaan door predatie en verstoring. Dit is echter geen reden tot paniek. Grutto’s zijn erop ingesteld dat hun broedsucces van jaar tot jaar varieert: mislukte jaren vormen geen probleem als er maar goede tegenover staan. In het Aubroek boden de kleine perceeltjes in beheer ook bescherming aan de grutto’s. Mochten deze perceeltjes er niet geweest zijn dan waren de resterende twee koppels ook uitgemaaid en had het BTS wellicht nog lager gelegen.
De gruttodichtheden in de Kalkense Meersen en het Aubroek liggen nog zeer laag in vergelijking met Nederlandse gebieden. In agrarisch grasland op Friese klei- en veengronden bedraagt de gemiddelde dichtheid 12 bp/km². In de reservaatgebieden wordt zelfs een gemiddelde dichtheid van 43 bp/km² waargenomen (Schekkerman et al. 2005). Mits een optimaal beheer is er dus nog heel wat potentieel om de gruttopopulatie uit te breiden.
5.1.2 Kievit
Het totaal aantal kievitkoppels in de reservaatzone en Broekmeers wordt op 55 geschat. Aangezien het studiegebied vrij groot is en omdat enkel vanaf de paden werd geïnventariseerd is dit mogelijk een onderschatting. Voor het vroegtijdig verlaten van de gruttoweide kunnen dezelfde oorzaken verantwoordelijk gesteld worden als bij de grutto’s. In het kerngebied op Belham deden de kieviten het beter en waren ze langer aanwezig dan de grutto’s. Er werden ook enkele vliegvlugge jongen gezien. Kieviten broeden in de Kalkensen Meersen ook op akkers en dit zorgt meestal voor nog meer problemen omdat op akkers geen bescherming wordt toegepast. Tijdens het voorjaar wordt een maïsakker minstens drie keer bewerkt (ploegen, rollen, inzaaien) waardoor kieviten telkens hun legsel verliezen en opnieuw moeten beginnen.
Ook in het Aubroek stelde dit probleem zich. Hier broedden minstens achttien koppels kievit waarvan zeker vijf op een akker. Met kieviten die daar in grasland broedden was het echter niet veel beter gesteld. Aangezien de meeste hooilanden in het Aubroek twee tot driemaal gemaaid worden tijdens het broedseizoen, al dan niet gevolgd door mestinjectie, moeten ze telkens vervolglegsels produceren. Gelukkig hebben kieviten een hoge capaciteit om meerdere legsels te leggen (Beintema et al. 1995). Toch is het opvolgen van het broedsucces van kieviten nodig om na te gaan of ze op deze manier wel nog genoeg jongen produceren om de populatie in stand te houden. Moest blijken dat de oudervogels elk jaar terugkomen en vervolglegsels produceren maar dat er niet voldoende jongen vliegvlug worden dan zou de populatie wel eens kunnen instorten op korte termijn.
5.2 Aandeel niet gemaaid grasland
Uit figuur 4.1 komt duidelijk naar voor dat in Broekmeers een groter aandeel niet gemaaid grasland aanwezig was tijdens de belangrijkste kuikenperiode (begin mei – half juni) in vergelijking met het Aubroek. In Broekmeers was gedurende de maand mei steeds minstens 40% (en max. 77%) meer ongemaaid grasland aanwezig dan in het Aubroek. Dit is hoofdzakelijk te danken aan de uitgestelde maaidata op de percelen die onder de beheersovereenkomsten van het weidevogelproject vallen. Een belangrijke opmerking hierbij is dat de grootte van dit verschil ook afhangt van het weer. Door het warme weer in het voorjaar van 2009 werd zeer snel gemaaid in het Aubroek. Bij slecht weer zullen percelen zonder beheersovereenkomsten ook later gemaaid worden en zal het verschil tussen ‘weidevogelpercelen’ en gangbare landbouwpercelen kleiner zijn, vooral in het begin van de kuikenperiode. Bovendien zal het verschil tussen de gebieden ook afhangen van het aantal ha dat onder weidevogelbeheer valt.
Bij uitbreiding van het weidevogelproject (wat het geval is in 2010) zal het verschil ook groter zijn en zal dit grotere verschil ook langer behouden blijven. De meerwaarde die een weidevogelproject biedt in vergelijking met gangbaar landbouwgebruik zal dus in grote mate afhangen van de omvang van het project en het weer.
In Broekmeers vonden de eerste maaibeurten twee weken later plaats dan in het Aubroek. Dit is positief voor de weidevogels, maar niet direct een gevolg van het weidevogelproject. Deze latere maaidata zijn eerder te wijten aan een tragere grasgroei in Broekmeers of hoogstwaarschijnlijk aan het feit dat in het Aubroek meer melkveehouders aanwezig zijn die nood hebben aan kuilgras. Kuilgras wordt namelijk vrij vroeg gemaaid.
In het Aubroek was bij het begin van de kuikenperiode slechts 12,7% van de oppervlakte grasland nog niet gemaaid t.o.v. 89,8% in Broekmeers. Deze 12,7% (6,4 ha) stelt niet veel meer voor en bestaat hoofdzakelijk uit de reservaatperceeltjes van vzw Durme. Als gevolg zijn de meeste nesten of kuikens die niet op een reservaatperceel broedden allicht verloren gegaan. Zelfs al overleefden enkele gruttokuikens, dan kwamen deze in een zee van ongeschikt habitat terecht. In gemaaid gras vinden gruttojongen niet veel voedsel en lopen ze een twee tot driemaal grotere kans om gepredeerd te worden (Schekkerman et al. 2005). Bovendien ontstaan serieuze afstanden tussen de resterende ongemaaide percelen. De vraag is of net uitgekomen gruttojongen überhaupt in staat zijn deze afstanden te overbruggen. Gruttogezinnen die door gebrek aan voldoende ongemaaid grasland gedwongen worden op gemaaide percelen te foerageren of er doorheen te trekken lopen dus een groter risico kuikens te verliezen. Daarenboven werden in het Aubroek verschillende percelen twee tot driemaal gemaaid wat wil zeggen dat vervolglegsels op die percelen ook al niet veel kans maakten.
Maaidata spelen dus een grote rol in het al dan niet behalen van voldoende broedsucces. Enerzijds heeft maaien een rechtstreekse invloed op de overleving van nesten en kuikens. Anderzijds heeft het een invloed op het voedselaanbod en foerageersucces van de gruttokuikens. Omdat gruttokuikens gemiddeld maar 26% van hun energie-inkomsten daadwerkelijk aan groei spenderen en nauwelijks langer kunnen foerageren dan de 11-14 uur per dag die ze er gemiddeld (in Nederlandse reservaten) al aan besteden, zal een afname in foerageersucces negatieve gevolgen hebben voor hun groeisnelheid en overleving (Schekkerman 2008). Uitstellen van de eerste snede is dus een belangrijke maatregel om de opgroeiomstandigheden voor gruttokuikens en de overleving van weidevogellegsels te verbeteren. Na midden juni heeft dit op bemeste percelen echter niet veel zin meer omdat de vegetatie dan te dicht wordt en het insectenaanbod afneemt (Schekkerman 2008). Maaien na 15 juni kan wel voordelig zijn voor laat broedende soorten zoals watersnip (Gallinago gallinago), zomertaling (Anas querquedula) en kwartelkoning (Crex crex) en soorten met tweede legsels (Oosterveld & Altenburg 2005).
Voor kievitkuikens is het negatieve effect op de voedselvoorziening minder van tel. Integendeel, vanaf midden mei worden ongemaaide percelen al onaantrekkelijk voor kievitkuikens door de hoge en dichte vegetatie. Kievitkuikens hebben namelijk een sterke voorkeur voor een korte vegetatie aangezien zij meer foerageren op insecten die op de bodem leven zoals kevers (Schekkerman 1997). De vochtigheidsgraad blijkt eveneens van belang. Op vochtige percelen worden meer potentiële prooidieren voor kievitkuikens gevonden dan op droge. Na maaien droogt het bodemoppervlak echter sneller uit waardoor het positieve effect voor kievitkuikens vermoedelijk van korte duur is (Schekkerman 1997). Kieviten zijn bijgevolg meer gebaat bij vochtige percelen met een (beperkt) aanbod van gemaaide percelen in de kuikenperiode tenzij er open plekken aanwezig zijn of een vegetatie voorkomt die van nature niet zo hoog en dicht is. In het noordelijk deel van Broekmeers is zo’n vochtig deel met een kortere vegetatie aanwezig wat het gebied aantrekkelijk maakt voor kieviten. Opvallend is dat het merendeel van de kievitkoppels ook daadwerkelijk op die plaats broeden. Hoewel kievitkuikens zich thuis voelen op gemaaide percelen is het toch belangrijk schuilmogelijkheden te voorzien zoals greppels of gemaaide perceelsranden.
Als besluit kan gesteld worden dat het weidevogelproject in 2009 effectief voor een hoger aandeel niet gemaaid grasland heeft gezorgd t.o.v. gangbaar landbouwgebied (het Aubroek). Toepassing van het weidevogelproject in zijn huidige vorm resulteert door de uitgestelde landbouwwerkzaamheden niet enkel in een reductie van nestverliezen maar ook in een hoger aandeel niet gemaaid grasland tijdens de belangrijkste kuikenperiode. Dit hoger aandeel ongemaaid grasland is één van de belangrijkste factoren die kan bijdragen tot een hoger broedsucces van gruttokoppels aangezien de overleving van jongen positief gecorreleerd is met het aandeel ongemaaid grasland (Schekkerman et al. 2005). Deze correlatie betekent echter ook dat maatregelen die specifiek op legseloverleving gericht zijn, zoals nestbescherming, in principe geen garantie bieden voor een voldoende broedsucces. Dit betekent niet dat nestgerichte maatregelen zinloos zijn, maar wel dat het rendement ervan aanzienlijk verhoogd kan worden door deze maatregelen te combineren met percelen met een latere maaidatum (Schekkerman 1997).
5.3 Het weer in 2009
Zoals eerder aangehaald (3.3) kan het weer op verschillende wijzen een invloed uitoefenen op het broedsucces van weidevogels. Ten eerste heeft het weer bij het begin van het broedseizoen een invloed op de legdatum. Als de start van het broedseizoen relatief warm is zal de legdatum vroeger liggen dan wanneer er nog nachtvorst aanwezig is. Het weer heeft verder ook een invloed op het verloop van agrarische werkzaamheden zoals het tijdstip waarop gemaaid wordt (en als gevolg het aanbod niet gemaaid grasland, zie 5.2) en het daaraan gekoppelde uitkomstsucces van legsels. Bovendien moeten eieren bij warm weer minder bebroed worden.
De belangrijkste invloed van het weer is echter die op de groei en overleving van kuikens. Grutto- en kievitkuikens zijn nestvlieders die vanaf het uitkomen zelf hun kostje bijeen moeten scharen. Aan deze levenswijze is een hoge energetische kost verbonden (40% hoger dan bij door hun ouders gevoerde nestblijvers, Schekkerman 2008) met als gevolg dat de kuikens het grootste deel van de daglichtperiode moeten besteden aan foerageren. Het foerageersucces wordt beïnvloed door het weer en het grootste effect is afkomstig van de wind. Vooral bij windsterktes boven 3-4 beaufort leidt de wind tot een afname van het foerageersucces. Wellicht omdat in het bewegende gras insecten minder zichtbaar en moeilijker te pakken zijn. Ook een toenemende temperatuur blijkt volgens Schekkerman (2008) het succes te verlagen, al kon dit niet onderscheiden worden van het tijdstip van de dag. Regen heeft ook een effect maar dit blijft eerder beperkt. Bij hevige regen stoppen kuikens echter volledig met foerageren. Regen en koud weer zijn zeer belangrijk in de eerste dagen na het uitkomen, dan moeten kuikens nog geregeld bebroed worden door de ouders. Hoe meer ze bebroed worden hoe minder tijd er overblijft om te foerageren. Op deze manier kunnen kuikens achterop geraken en kuikens met een lagere conditie hebben een hogere kans om te verdwijnen, enerzijds door predatie, anderzijds door uitputting (Schekkerman et al. 2005).
In tegenstelling tot gruttokuikens die overdag nauwelijks meer bebroed worden na tien dagen, worden kievitkuikens zelfs na drie weken nog bebroed. Dit is te wijten aan een langzamer ontwikkelend metabolisme bij kievitkuikens (Visser & Ricklefs 1993). Het resultaat is dat kievitkuikens gemiddeld minder tijd kunnen besteden aan foerageren en dat ze hierdoor gevoeliger zijn voor weersinvloeden dan grutto’s (Schekkerman 1997).
Behalve deze rechtstreekse weersinvloeden zijn er ook interacties mogelijk tussen het graslandbeheer en het weer op het insectenaanbod. Bij het ouder worden hebben de gruttokuikens grotere prooien nodig om aan hun energiebehoefte te voldoen. Als grote insecten minder aanwezig zijn als gevolg van het beheer (lees: een hoge bemesting, zie ook 5.7) zullen ze meer tijd moeten besteden aan foerageren en zijn ze gevoeliger voor weersinvloeden.
Tenslotte heeft het weer, in combinatie met de grondwaterstand, een invloed op de voedselbeschikbaarheid voor volwassen vogels. Bij droog weer zullen regenwormen eerder dieper in de bodem trekken en zal minder voedsel beschikbaar zijn voor volwassen vogels. Bovendien wordt de indringingsweerstand van de bodem dan snel te groot waardoor grutto’s niet meer in de bodem kunnen foerageren. De vochtigheidsgraad blijkt, behalve voor volwassen vogels, ook belangrijk voor de voedselbeschikbaarheid van kievitkuikens (Schekkerman 1997).
2009 werd gekenmerkt door een zeer warm en vrij droog voorjaar. Dit leidde enerzijds tot snelle maaidata en anderzijds tot een snelle uitdroging van de graslanden in de deelgebieden van deze studie. In het Aubroek resulteerde dit weer in slechte omstandigheden voor zowel de kuikens als voor volwassen vogels. In Broekmeers werden de maaiverliezen beperkt door de uitvoering van het weidevogelproject en bleef bovendien een groter aandeel kuikenland over. In combinatie met het voor het foerageersucces gunstige weer is dit waarschijnlijk de verklaring voor de hoge BTS-waarde die gevonden werd in Broekmeers. Het weidevogelproject speelt via het uitstellen van de eerste snede dus ook in op de weersinvloeden, vooral bij warm en droog weer.
5.4 Historisch verloop gruttopopulatie
Eerst en vooral moet erop gewezen worden dat de oudste indicatie voor het voorkomen van grutto’s misschien wel met een korreltje zout moet genomen worden. Volgens van Damme & Lostrie (1987) waren grutto’s vroeger zo talrijk aanwezig in de regio dat hun eieren verzameld en verkocht werden voor de tafels der rijken. Er is wel niet nader gespecificeerd wat bedoeld wordt met vroeger. Als men echter bij de oude landbouwers navraag doet over het voorkomen van grutto’s in de Kalkense Meersen herinneren zij zich niet dat de grutto vroeger (jaren ’30 - ’50) voorkwam en blijkt ‘de koning der weidevogels’ hier misschien toch een ‘recente’ verschijning sinds de jaren ‘60.
Het is wel mogelijk dat grutto’s een paar eeuwen geleden algemeen waren en daarna verdwenen, maar hiervoor werden geen bewijzen gevonden.
Volgens de broedgegevens broedde de grutto in de beginjaren (jaren ’60 en ’70) in Wijmeers. Zij waren daar gedurende enkele jaren aanwezig met twee à vier koppels. In 1979 vond een verschuiving plaats van één koppel naar Langendonk en tussen 1980 en 1982 een verdere verschuiving van de koppels van Wijmeers naar Langendonk, Molenmeers en Broekmeers. Volgens Caekebeke & De Bruycker (1984) is deze verschuiving te wijten aan een daling van het waterpeil als gevolg van het uitbaggeren van de grachten in Wijmeers in het voorjaar van 1979. Nadien werd, tot in 2008, in Wijmeers geen enkel koppel meer waargenomen. De twee knikken in de stijgende groeicurve van de gruttopopulatie in 1993 en 1997 (fig. 4.2) zijn volgens sommige historische gegevens te wijten aan weersomstandigheden. Voor het lagere aantal broedparen in 1997 word het droge voorjaar van het jaar voordien als mogelijke oorzaak vooropgesteld (geg. R. Clinckspoor). Tevens wordt de extreem droge en strenge winter aangehaald (Claus 2005). Op zich heeft een strenge winter niet veel effect op de vogels zelf aangezien ze in Afrika overwinteren. Een mogelijk effect van een strenge winter kan zich wel uiten in de abundantie van regenwormen, het stapelvoedsel van volwassen grutto’s. Timmerman et al. (2006) constateerden dat koude winters resulteren in een lagere abundantie van regenwormen in de komende lente. Of dit één van de redenen is valt natuurlijk niet met zekerheid te zeggen. Het jaar nadien (1998) was er alleszins niet veel meer van te merken en steeg de populatie verder tot een maximum van 30 koppels in 2002.
De laatste jaren blijft de populatie in de Kalkense Meersen hangen op 25 koppels. Als oorzaak van de daling in 2003 worden in de historische gegevens dezelfde redenen aangehaald als bij de lichte knikken van 1993 en 1997. Opvallend is wel dat deze stagnatie ongeveer samenvalt met het verschijnen van broedende grutto’s in andere gebieden in de regio. Rond het station van Schellebelle vestigden zich al broedende grutto’s in 1998 maar in het Aubroek (Berlare) en de Paardenweide (Wichelen) werden ze voor het eerst waargenomen vanaf 2001. In deze omliggende gebieden is geen afname te zien na 2002. Integendeel, er wordt zelfs een toename waargenomen (fig. 4.3). In 2005 broedden er bijna dubbel zoveel koppels als in 2002. Deze trend kan op drie verschillende manieren verklaard worden. Ten eerste kan het erop wijzen dat enkele koppels om een of andere reden verhuisd zijn van de Kalkense Meersen naar een ander gebied in de regio. Het is ook mogelijk dat de stijging van het aantal koppels in de regio verder ging in de jaren 2003 – 2005 maar dat de nieuwe koppels eerder de randgebieden verkozen dan de Kalkense Meersen zelf. De derde mogelijke verklaring ligt in het feit dat de toename in de randgebieden te wijten is aan jongenaanwas. Dit impliceert echter dat er in de Kalkense Meersen de laatste jaren geen jongenaanwas plaatsvond. Claus (2005) haalt aan dat in de periode 2003 – 2005 waarschijnlijk een laag broedsucces behaald is door het vroegtijdig maaien, wat een mogelijke verklaring zou kunnen zijn voor een beperkte aanwas van jongen in de daaropvolgende jaren.
De echte oorzaak kan ook uit een combinatie van bovenstaande verklaringen bestaan. Hoe dan ook suggereren deze verklaringen dat de Kalkense Meersen op één of andere wijze niet meer geschikt lijken voor nieuwe koppels om zich te vestigen en te broeden en/of om voldoende jongen groot te brengen. In die zin is het koppel dat sinds een aantal jaar op de Molenveldkouter broedt veelzeggend. Dit koppel broedt in een raaigrasperceel dat volledig omgeven is door zandige akkers, wat zeker geen optimaal habitat is, integendeel. Het valt wel niet uit te sluiten dat we hier te maken hebben met een afwijkend geval.
Betekenen deze cijfers dan dat de gruttopopulatie zijn draagkracht bereikt heeft in de Kalkense Meersen? Dit lijkt onwaarschijnlijk aangezien het gebied vrij groot is en er nog geschikt habitat voor grutto’s onbezet is. Bovendien ligt de gruttodichtheid in de Kalkense Meersen nog zeer laag in vergelijking met de gemiddelde dichtheden in Nederland (vergelijk met Schekkerman et al. 2005). De ongeschiktheid als broedhabitat uit zich waarschijnlijk eerder in een combinatie van factoren. Enerzijds zorgen de bosjes, knotbomen en andere kleine landschapselementen (KLE’s) ervoor dat de reservaatzone niet open genoeg is en dat hiermee een hogere predatiedruk gepaard gaat. Anderzijds wordt Broekmeers, de enige echte grote, open grasvlakte in de meersen, redelijk intensief bewerkt waardoor grote verliezen onder eieren en kuikens optreden. Dit kan leiden tot een zeer laag broedsucces. De randgebieden worden trouwens ook nog intensief bewerkt waardoor in die gebieden ook niet zo’n hoog broedsucces verwacht wordt. Ondanks het feit dat de broedplaatstrouw bij grutto’s gekoppeld is aan het broedsucces (Schekkerman et al. 2005), blijven de 25 koppels toch aanwezig in de Kalkense Meersen. Over de jaren heen zijn er evenwel grote verschuivingen waar te nemen in de kerngebieden van de grutto’s.
Volgend op de verschuiving van Wijmeers naar Langendonk, Molenmeers en Broekmeers werden in de periode 1985 - 1995 alle deelgebieden van het studiegebied stilaan gekoloniseerd door de groeiende gruttopopulatie (fig. 4.4). Als resultaat was elk deelgebied in 1996 bezet met een viertal koppels. Enkel Scherenmeersen had er toen zeven. Nadien steeg het aantal koppels in Molenmeers opvallend sterk tot een maximum van tien koppels in 2000. Ook het aandeel van Belham steeg toen een beetje. Na deze piekjaren ging het echter snel bergaf met Molenmeers en Scherenmeers. Molenmeers verloor waarschijnlijk het merendeel van zijn koppels aan Broekmeers (fig. 4.5). De koppels van Scherenmeersen verhuisden allemaal, waarschijnlijk naar de aangrenzende deelgebieden Belham en Langendonk (fig. 4.6). Er moet wel vermeld worden dat de grenzen tussen de deelgebieden, vooral tussen Langendonk en Scherenmeersen, door de mens bepaald zijn en geen echte fysieke barrière vormen. De grenzen vallen meestal wel samen met waterlopen zoals grote beken, maar voor grutto’s vormen deze geen obstakel. Daarom kunnen verschuivingen zoals die tussen Langendonk en Scherenmeersen in 1988 – 1989 puur op toeval gebaseerd zijn omdat enkele koppels bijvoorbeeld een tiental meter opgeschoven zijn. De laatste jaren (2000-2009) zijn de verschuivingen daarentegen veel duidelijker en gaan ze over grotere afstanden.
Naast verschuivingen tussen deelgebieden vonden ook duidelijke verschuivingen binnen deelgebieden plaats. Dit fenomeen heeft zich voornamelijk voorgedaan in Langendonk en Belham in de periode 2000 – 2002. Vóór 2000 broedden de grutto’s van Langendonk bijna allemaal in het noordoostelijk deel. Tussen 2000 en 2002 zijn alle koppels de Blokstraat overgestoken naar de gruttoweide, waar ze ook gebleven zijn tot en met 2009. In Belham broedden de koppels vóór 2000 vooral in het noordelijk deel, tegen Scherenmeers. Vanaf 2002 zitten ze bijna allemaal zuidelijker tegen de Belhamstraat aan (Zijmeers).
De achterliggende oorzaken van deze verschuivingen, zowel tussen als binnen deelgebieden, zijn niet altijd eenduidig en het is bijgevolg geen sinecure om ze te proberen verklaren. In wat hier volgt is toch getracht enkele factoren aan te duiden die (mede) bepalend kunnen geweest zijn voor dergelijke verschuivingen.
Het eerste grote punt dat belangrijk kan zijn is het beheer. Het natuurbeheer dat in de Kalkense Meersen gevoerd wordt d.m.v. hooien en nulbemesting leidt vooral tot een verschraling en soortenrijkere plantengemeenschappen. Deze vegetatie is echter minder fors en is meestal nog niet erg hoog opgeschoten wanneer de grutto’s aankomen. Bovendien zijn in bemeste bodems dikwijls meer regenwormen, het stapelvoedsel van volwassen grutto’s, aanwezig (o.a. Standen 1984). Mogelijk leidt dit natuurbeheer dus tot graslanden die niet meer geschikt zijn voor volwassen grutto’s aangezien zij lang gras verkiezen om een nest in te bouwen en omdat er op de bemeste percelen meer voedsel aanwezig is. De verschuiving van Molenmeers naar Broekmeers en de verschuiving die plaats vond binnen Langendonk (van het NO deel naar de gruttoweide) zijn hier mogelijk door te verklaren. In Molenmeers en het NO deel van Langendonk heeft Natuurpunt namelijk een groot deel van de percelen in beheer sinds het begin van de jaren ’90. Deze hypothese was de aanzet tot verder onderzoek waarin onderzocht werd of niet bemeste percelen van Molenmeers werkelijk minder wormen bevatten en of de vegetatie er ook korter was dan op bemeste percelen van Broekmeers (dit wordt verder uitgediept in secties 5.5 en 5.6).
Een tweede factor die aan de basis kan liggen van een verschuiving is verstoring. Verstoring kan zich voordoen onder allerlei vormen zoals predatie, verkeer, landbouwwerkzaamheden, aanwezigheid van andere organismen die het habitat ongeschikt maken, enzovoort. Verstoring onder de vorm van predatie en verkeer werd reeds besproken in 5.1.1. De gevolgen van landbewerkingen zoals bemesten en maaien tijdens het broedseizoen spreken voor zich. Vele nesten en jongen overleven het niet waardoor de vogels een laag broedsucces behalen. Bij grutto’s is de broedplaatstrouw gekoppeld aan het broedsucces (Schekkerman et al. 2005). Dit wil zeggen dat koppels normaal gezien elk jaar naar dezelfde plaats terugkeren om te broeden als op die plaats voldoende broedsucces behaald werd in de voorgaande jaren. Bij een laag broedsucces is het waarschijnlijk dat ze betere oorden opzoeken. In ons geval zal dit leiden tot een verschuiving, ofwel binnen de Kalkense Meersen, ofwel naar andere gebieden. Als laatste vorm van verstoring wil ik wijzen op andere organismen die door hun aanwezigheid en levenswijze een negatief effect kunnen hebben op de grutto’s. Een voorbeeld hiervan zijn de ganzen die sinds een tiental jaar talrijk aanwezig zijn in de Kalkense Meersen. Deze ganzen (vooral Canadese ganzen (Branta canadensis) en Nijlganzen (Alopochen aegyptiaca)) zijn het hele jaar door aanwezig op Scherenmeersen, exact op dezelfde percelen waar vroeger het merendeel van de grutto’s broedde. Enerzijds zorgen deze relatief agressieve en luidruchtige ganzen voor verstoring door hun talrijke aanwezigheid. Anderzijds grazen ze het gras kort en deponeren ze de percelen vol met (zure) uitwerpselen waardoor deze niet meer geschikt zijn als broedhabitat voor grutto’s. De aanwezigheid van deze ganzen is waarschijnlijk één van de verklaringen waarom Scherenmeersen de laatste jaren zijn status als kerngebied verloren is.
De verschuiving binnen Belham is mogelijk te wijten aan het feit dat de percelen langs de Belhamstraat natter zijn waardoor ze later gemaaid worden. Bovendien is daar meer microreliëf aanwezig onder de vorm van depressies, bultjes en ondiepe beekjes wat bevorderlijk is voor weidevogels. Microreliëf zorgt voor afwisseling in de vegetatie waardoor ze geschikt wordt als broedhabitat voor verschillende soorten weidevogels. Als gevolg hiervan kunnen hogere dichtheden bereikt worden. Weidevogels zijn ervoor gekend dat ze samenwerken bij het verdedigen van nesten en jongen tegen predators en hogere dichtheden leiden bijgevolg tot een hogere efficiëntie van deze verdedigingstechniek. Deze efficiëntere verdediging kan ook verklaren waarom de grutto’s enkel in een paar clusters voorkomen binnen de Kalkense Meersen en dat weinig koppels apart broeden. Het is te vergelijken met het zogenaamde ‘Allee-effect’ (o.a Stephens 1999). Dit is het effect waarbij kleine populaties van organismen ineenstorten als de populatiegrootte onder een bepaalde drempelwaarde zakt. Dit effect treedt bijvoorbeeld op bij sociale organismen waarbij onder een bepaalde populatiegrootte de sociale structuur van de populatie uiteen valt.
In het geval van de grutto’s is de efficiëntie van het verdedigen veel lager bij lage dichtheden. Ze moeten alle verdediging zelf doen en verliezen daarbij ook veel meer energie. Als gevolg zal het broedsucces in de meeste gevallen laag zijn en kan de populatie op termijn ineenstorten. Verschuivingen van alle koppels uit een bepaald gebied naar een ander gebied kunnen dus ook het gevolg zijn van een paar koppels die verhuizen. De andere koppels volgen dan mogelijk om te kunnen blijven profiteren van de hogere dichtheid.
Voor de kievit was het niet mogelijk een historisch verloop te reconstrueren omdat er niet genoeg gegevens beschikbaar waren. Kievitpopulaties zijn in het verleden nooit echt goed geïnventariseerd geweest. Op telformulieren van broedvogelinventarisaties wordt meestal enkel aangeduid dat de soort aanwezig is maar het aantal broedparen wordt niet geteld of vermeld. Het opvolgen van de kievitpopulaties is echter nodig om een achteruitgang van deze soort tijdig op te merken. Kievitpopulaties hebben tegenwoordig ook veel te lijden onder de intensivering van de landbouw en zouden, zoals in 5.1.2 gesteld, wel eens kunnen instorten in de nabije toekomst.
5.5 Voedselaanbod onder de vorm van regenwormen
Uit het historisch overzicht van de broedlocaties van grutto’s in de Kalkense Meersen (5.4) blijkt de laatste jaren een duidelijke verschuiving aan de gang te zijn. Sinds de eeuwwisseling zijn grutto’s deelgebieden waar tegenwoordig een groot aandeel van de percelen onder natuurbeheer valt beginnen mijden en trekken ze naar meer intensievere deelgebieden die nog bemest worden. Deze duidelijke verschuiving riep nieuwe vragen op waarvan de antwoorden mogelijk een verklaring bieden voor deze veranderde nestplaatskeuze. Is er op bemeste percelen meer voedsel aanwezig voor volwassen grutto’s onder de vorm van regenwormen? Is de vegetatie hoger op bemeste percelen op het moment dat ze aan een nest beginnen? Hoe zit het met de kruidenrijkdom op de bemeste percelen en wat is het gevolg daarvan op de voedselbeschikbaarheid voor kuikens? Het antwoord op de twee laatste vragen wordt respectievelijk gegeven in secties 5.6 en 5.7. In deze sectie wordt ingegaan op het verschil in het aantal en de biomassa regenwormen tussen bemeste percelen van Broekmeers en onbemeste percelen van Molenmeers. Er wordt ook aandacht besteed aan de algemene effecten van bemesting en de grondwaterstand op de bodemfauna en verschillen tussen bemestingsvormen aan de hand van wat in de huidige beschikbare literatuur te vinden is.
In de proefvlakken op bemeste percelen van Broekmeers werden bijna driemaal zoveel regenwormen m-2 gevonden dan in de proefvlakken op niet bemeste percelen in Molenmeers (fig. 4.7). De aantallen die in de resultaten (zie 4.4) weergegeven worden zijn mogelijk nog een onderschatting van het werkelijk aantal wormen dat aanwezig is omdat bij het uitsorteren maar de bovenste 15 cm van de bodem opgegraven werd. Wormen zitten dikwijls ook dieper, afhankelijk van de grondwaterstand. Toch is het gevonden aantal wormen representatief voor het voedselaanbod voor grutto’s aangezien ze met hun snavel enkel de bovenste 12 cm kunnen bereiken. Ook de totale biomassa van regenwormen lag veel hoger in bemeste proefvlakken (fig. 4.8). De verklaring hiervoor ligt hoofdzakelijk bij het grotere aantal regenwormen in bemeste proefvlakken en niet omdat de wormen daar ook zwaarder wogen. De gemiddelde massa van wormen was wel iets hoger in de bemeste proefvlakken, maar dat verschil was niet significant. De zwaarste wormen (>1g) werden wel bijna allemaal gevonden in bemeste proefvlakken maar aangezien het verschil in gemiddelde gewicht niet significant is wijst dit erop dat er in vergelijking met onbemeste proefvlakken ook zeer veel lichte wormpjes aanwezig waren. De gevonden biomassa van 159,4 g m-2 is hoog (Timmerman et al. 2006). In goed bemeste weidevogelreservaten wordt 70 – 120 g m-2 gevonden in de periode april/mei (Brandsma 1999). In weidevogelreservaten wordt meestal wel iets minder bemest dan op gangbaar agrarisch grasland zoals Broekmeers. De kritische ondergrens ligt voor de grutto bij 15 – 30 gram regenwormen m-2 en de aantallen nemen doorgaans boven deze grens al af (Brandsma 1999). Volgens Hut & Helmig (2003) ligt de grens tussen goede en minder goede weidevogelreservaten rond 60 g m-2. Het is duidelijk dat Molenmeers met gemiddeld 35,7 gram m-2 deze grens op het moment van de bemonstering niet haalt.
Of dit duidelijke verschil in aantal en biomassa enkel te wijten is aan een verschil in bemesting is niet eenduidig. Hoogstwaarschijnlijk speelt er ook een effect van de grondwaterstand mee. Hoewel de locatie van de proefvlakken in bemeste en onbemeste percelen bepaald werd op basis van een gelijkaardige bodemtextuur en hydrologie waren de percelen in Molenmeers nog veel natter op het moment van de staalname. Bij langdurige overstroming is bekend dat regenwormpopulaties sterk afnemen of zelfs verdwijnen (o.a. Plum 2005). Ausden et al. (2001) stelden vast dat de biomassa van de meeste bodembewonende macroinvertebraten sterk afnam in gebieden die onderhevig zijn aan overstroming tijdens de winter, ondanks het feit dat verschillende soorten regenwormen lange perioden van overstroming kunnen overleven. Men veronderstelt dat regenwormen overstroomde bodems vermijden wegens een gebrek aan zuurstof en het voorkomen van schadelijke gassen, eens anoxische condities optreden (Mather & Christensen 1988). Bovendien lag in de studie van Ausden et al. (2001) de biomassa van macroinvertebraten veel lager op recent overstroomde percelen in vergelijking met percelen die voorheen al meerdere jaren geconfronteerd werden met winteroverstroming. De auteurs schrijven dit effect toe aan de grotere hoeveelheid afbrekend organisch materiaal dat dikwijls aanwezig is op recent overstroomde percelen. Grote hoeveelheden afbrekend materiaal consumeren immers sneller de beschikbare zuurstof en resulteren bijgevolg snel in anoxische condities.
Regenwormen zijn in staat om bovengronds relatief grote afstanden te overbruggen (Mather & Christensen 1992, Darwin 1881). Volgens Ausden et al. (2001) is de verklaring voor de lagere biomassa in overstroomde percelen dan ook te zoeken in het feit dat ze de bodem verlaten wanneer deze onder water komt te staan. Deze regenwormen zullen zich dan concentreren in de bovenste lagen van niet overstroomde bodem, wat tevens de aantrekking van steltlopers kan verklaren aan de rand van overstroomde graslanden. Verder was er tijdens de studie niet echt sprake van een rekolonisatie tijdens de lente, wat volgens de auteurs niet verwonderlijk is aangezien sommige soorten vrij trage kolonisatiesnelheden hebben. Bovendien wordt rekolonisatie bemoeilijkt door effecten van de overstroming zelf. Overstroming kan leiden tot een compactere bodem en het verdwijnen van een duidelijke bodemstructuur, wat lucht- en waterbewegingen moeilijker maakt en bijgevolg ook de voortbeweging van regenwormen belemmert.
Het grondwater staat in het gros van de niet bemeste percelen in Molenmeers tijdens de winter boven het maaiveld. Het is dus waarschijnlijk dat de grondwaterstand een grote rol speelt in de verschillen die gevonden werden tussen Molenmeers en Broekmeers. Omdat Molenmeers iets lager ligt dan het grootste deel van Broekmeers zal het waterpeil daar later in het voorjaar onder het maaiveld zakken en kunnen de regenwormen bijgevolg de bodem van Molenmeers maar later rekoloniseren. Dit kan ook verklaren waarom in de niet bemeste proefvlakken zeer veel kleine, witte wormpjes gevonden werden, waarschijnlijk juvenielen, wat kan wijzen op het begin van de rekolonisatie. Omdat ik geen kennis heb van de verschillende soorten wormen valt wel niet uit te sluiten dat dit gewoon een andere soort was. Deze minieme wormpjes werden niet meegeteld omdat ze zo klein zijn dat ze vermoedelijk geen bijdrage leveren aan het voedsel van grutto’s.
De rol die de bemesting speelt in het verschil is eveneens niet zo eenvoudig vast te stellen omdat factoren zoals het weer, bodemtype en vegetatie ook een invloed uitoefenen op regenwormpopulaties. In het algemeen neemt het aantal en de biomassa regenwormen toe met een toenemende bemesting (o.a. Hansen & Engelstad 1999), en zijn deze parameters positief gecorreleerd met de pH (Standen 1984, Oosterveld & Altenburg 2005). Als de bodem pH onder 4,5 zakt zijn er meestal geen regenwormen meer aanwezig (Curry 2004). Bemesting met organische mest zorgt voor een toename van organisch materiaal (het voedsel van regenwormen) en houdt de pH op peil (Oosterveld 2006). Uit enkele studies blijkt de pH echter een belangrijkere factor dan bemesting. Verhulst et al. (2007) geven aan dat een toename van de prooidensiteit niet altijd verwezenlijkt kan worden door een hogere mestgift maar dat eerder de nadruk moet gelegd worden op het behouden van een hoge pH. De verrassendste resultaten zijn afkomstig van Timmerman et al. (2006). Deze onderzoekers constateerden dat na 25 jaar zonder bemesting het aantal wormen niet daalde en dat er zelfs geen verschil aanwezig was in vergelijking met bemeste percelen. Het aantal wormen was zelfs hoger in de onbemeste behandeling dan in de behandeling met drijfmest. Deze bevindingen zijn dus totaal tegengesteld aan de algemene gedachte, dat de abundantie van regenwormen stijgt bij bemesting (Curry 2004).
De auteurs suggereren dat deze resultaten gerelateerd zijn aan de hydrologie en bodemcondities van het onderzoeksgebied. Het gebied (in Nederland) wordt gekenmerkt door een hoge grondwatertafel en de uittredende kwel resulteert in een relatief hoge natuurlijke bodem pH. Door het optreden van kwel tot in het maaiveld is dus ook zonder bemesting en bekalking de zuurtegraad en de hoeveelheid organisch materiaal blijkbaar geschikt voor het talrijk voorkomen van regenwormen (Oosterveld 2006).
Aangezien in Molenmeers ook kwel optreedt tot in het maaiveld en het bodemtype gelijkaardig is aan dat uit de studie van Timmerman et al. (2006) (klei op veen), kan verwacht worden dat Molenmeers ook aanzienlijke densiteiten aan regenwormen kan bevatten. Waarschijnlijk treden deze grotere aantallen pas later op in het voorjaar, na de rekolonisatie die mogelijk gemaakt wordt door een daling van de grondwaterstand. Dit kan echter pas geverifieerd worden wanneer de bodem in Molenmeers bemonsterd wordt op een later tijdstip in het voorjaar, wat een suggestie kan zijn voor verder onderzoek.
Naast de hoeveelheid toegediende mest heeft ook de bemestingsvorm een invloed op het aantal en de biomassa regenwormen. Over het algemeen wordt best organische mest gebruikt omdat deze de beste resultaten levert.
In de meeste studies levert organische mest een hogere abundantie van regenwormen op terwijl de effecten van anorganische mest (minerale mest) variëren (Estevez et al. 1996). De negatieve effecten van anorganische meststoffen zijn veelal te wijten aan verzuring van de bodem door bijvoorbeeld ammonium of sulfaat (Ma et al. 1990). Bovendien voegt kunstmest geen verse organische stof toe aan de bodem (Oosterveld 2006).
Bij bemesting met organische mest is ook het soort mest (drijfmest, vaste mest) en de wijze waarop deze toegediend wordt van belang. Vaste mest blijkt in de meeste studies de beste resultaten op te leveren (o.a. Oosterveld & Altenburg 2005, Timmerman et al. 2006, Oosterveld 2006). Na toediening van drijfmest is soms een negatief effect waar te nemen op de regenwormenpopulaties. Oosterveld (2006) constateerde dat in de eerste vier weken na de toediening van drijfmest de biomassa aan regenwormen afnam met 54% (zodenbemesting: hierbij wordt mest in geultjes van 5 cm diep in de bodem gebracht) en 60% (sleepvoetenbemesting: hierbij wordt mest in stroken op en tussen het gras gelegd) en bij vaste mest gelijk bleef. Na vier weken was dit verschil wel verdwenen. Verder werd bij zodenbemesting en sleepvoetenbemesting respectievelijk 35% en 25% lagere aantallen wormen gevonden in vergelijking met vaste mest. Tussen de biomassa werd echter geen verschil gevonden wat erop wijst dat bij drijfmest minder, maar zwaardere wormen aanwezig waren. De invloed van zodenbemesting is vrij complex. Soms resulteert zodenbemesting in een afname, soms in een toename van het aantal regenwormen. In het geval van een afname is dit meestal van tijdelijke aard en treedt binnen één tot enkele maanden herstel op (Huijsmans et al. 2008). Deze afname gedurende een maand of langer kan wel van belang zijn aangezien meestal bemest wordt in maart, wanneer de grutto’s net toekomen. In de studie van Oosterveld (2006) werden evenwel geen bewijzen gevonden voor een lager foerageersucces na zo’n afname. Grutto’s blijken bij 100 gram regenwormen m-2 genoeg energie te kunnen verzamelen als ze 5,5 uur foerageren per dag (Wymenga & Alma 1998).
Aangezien bij vaste mest geen incidentele afname geconstateerd wordt en dat in het algemeen betere resultaten bereikt worden dan met drijfmest (Timmerman et al 2006, Oosterveld 2006, Hansen & Engelstad 1999), is de algemene gedachte, dat vaste mest de beste oplossing biedt in weidevogelreservaten, waarschijnlijk wel gerechtvaardigd. Bovendien kan vaste mest volgens Timmerman et al. (2006) nog een meerwaarde bieden als nestmateriaal voor vogels of via effecten op de bodemstructuur en de entomofauna. Ondanks de aanbeveling voor vaste mest, worden met drijfmest ook nog zeer goede resultaten bereikt. Timmerman et al. (2006) zien dan ook geen echte reden om drijfmest te bannen en Oosterveld (2006) hanteert als vuistregel dat men beter drijfmest gebruikt dan geen mest. Oosterveld (2006) vertelt er ook wel bij dat deze vuistregel niet overal geldt. Zoals blijkt uit de studie van Timmerman et al. (2006) moet men in gebieden met kwel tot in het maaiveld terughoudend zijn met drijfmest. Toepassing van drijfmest leverde daar immers 20-30% minder regenwormen op dan bij vaste mest en geen bemesting !
Om te bemesten moeten landbouwers natuurlijk hun land op. De landbouwwerkzaamheden zelf hebben volgens Hansen & Engelstad (1999) ook nog een effect op het aantal regenwormen. In hun studie werden minder wormen gevonden wanneer de intensiteit van het ‘landbouwverkeer’ hoog was.
Uit al deze informatie over de invloed van de grondwaterstand en bemesting op het voorkomen van regenwormen kan afgeleid worden dat de oorzaak van het grote verschil tussen Molenmeers en Broekmeers vooral te zoeken is bij de hogere grondwaterstand. Het tijdstip waarop dit grote verschil optreedt is wellicht afhankelijk van de weersomstandigheden. Februari en maart waren in 2010 vrij koud. Als het weer in deze maanden redelijk droog en warm is zal in Molenmeers het grondwaterpeil sneller dalen en zal de regenwormenpopulatie zich sneller kunnen herstellen. Daartegenover staat dat de wormen op de iets drogere percelen van Broekmeers evenwel sneller in het voorjaar diepere grondlagen zullen opzoeken waardoor de voedselbeschikbaarheid voor grutto’s daar lager kan uitvallen. De ongeschiktheid van Molenmeers als foerageergebied voor volwassen vogels bij de start van het voorjaar zal dus variëren met de jaren. Door de hogere watertafel wordt Molenmeers waarschijnlijk wel belangrijker als foerageergebied naarmate het broedseizoen vordert en de percelen van Broekmeers te droog worden. De staalname in het kader van deze studie is vermoedelijk wel niet 100% representatief voor heel Broekmeers maar eerder voor de iets hoger gelegen percelen aan de oostelijke zijde. Het noordwestelijke deel van Broekmeers is lager gelegen en daar staat het water in de winter en het voorjaar ook boven het maaiveld. De situatie zal daar dus vermoedelijk gelijkaardig zijn aan die van Molenmeers.
Om de bemestingsstrategie voor een weidevogelreservaat te kiezen is het van belang zowel de zuurtegraad als de bodemfauna te kennen. Een eenmalige bemonstering van de bodemfauna is daarbij onvoldoende omdat de weersomstandigheden het effect van bemesting kunnen overheersen. Timmerman et al. (2006) constateerden namelijk dat er een grote jaarlijkse variatie optreedt in aantallen en biomassa regenwormen die het gevolg is van hoe streng de voorafgaande winter is. Na koude en natte winters worden in het voorjaar minder regenwormen aangetroffen. Deze grote variatie tussen jaren benadrukt de voorzichtigheid waarmee resultaten van korte termijn studies met een beperkt aantal proefvlakken, zoals deze, geïnterpreteerd moeten worden.
Aangezien de bodem pH in Molenmeers en Broekmeers optimaal is voor het in stand houden van aanzienlijke regenwormpopulaties zou ik aanraden terughoudend te zijn met bemesting voor het weidevogelgebied. Bemesting houdt de grasgroei op peil maar hypothekeert meestal het voedselaanbod voor de kuikens (zie verder, 5.7). Verder onderzoek naar de pH en de bodemfauna bij afwezigheid van bemesting is daarom gewenst in het toekomstige weidevogelgebied van het Sigmaplan.
5.6 Vegetatiehoogte
Naast het voedselaanbod kan ook de hoogte van de vegetatie een rol spelen in de nestplaatskeuze van grutto’s. Grutto’s maken hun nest immers liefst in vrij hoog gras (Beintema et al. 1995). Enerzijds houdt bemesting de grasgroei op peil (Oosterveld 2006). Anderzijds zorgt een verhoging van de watertafel, met overstroming tijdens de winter als gevolg, voor een lagere vegetatie in het voorjaar (Ausden et al. 2001). Om deze redenen zal de vegetatie op niet bemeste percelen met een hogere waterstand in het voorjaar lager zijn dan op bemeste, drogere percelen. Verschraling en het verhogen van de watertafel zijn nu net de kenmerken van het natuurbeheer dat in het grootste deel van Molenmeers ingevoerd wordt sinds de jaren ‘90. Het natuurbeheer resulteert dan ook in een lagere vegetatie bij de start van het broedseizoen.
Eind maart, begin april was de vegetatie in Broekmeers gemiddeld ongeveer 1 cm hoger dan in Molenmeers, wat eigenlijk niet zo veel is (zie fig. 4.9). Het verschil ligt vermoedelijk hoger omdat op de percelen in Molenmeers veel afgestorven, harde plantendelen aanwezig waren van het vorige seizoen. Het plastieken mapje bleef eerder rusten op deze afgestorven plantendelen dan op de verse, groene vegetatie. Dit heeft geleid tot een lichte overschatting van de gemiddelde vegetatiehoogte in Molenmeers. Bovendien was een week later de hoogte van de vegetatie in Broekmeers bijna verdubbeld terwijl die in Molenmeers nog zeer laag bleef (eigen waarneming). Kleijn et al. (2007) constateerden dat de hoogteontwikkeling 2-3 keer zo snel gaat op gangbaar beheerde percelen (veel mest) in vergelijking met kruidenrijke percelen (beperkte mestgift).
Deze waarnemingen bevestigen dat in Molenmeers (en ook in Langendonk) de vegetatie van verschraalde percelen bij de start van het broedseizoen minder geschikt is voor de grutto’s om in te nestelen. Het is daarom zeer waarschijnlijk dat deze factor mee bepalend is voor de verschuiving naar Broekmeers die de laatste jaren aan de gang is. Dit neemt niet weg dat een korte vegetatie niet gunstig is voor andere weidevogels. Kieviten, en ook tureluurs (Tringa totanus) hebben een voorkeur voor een kortere vegetatie. De vegetatie die ontstaat bij natuurbeheer zal voor deze soorten dus betere nestgelegenheden bieden dan hoog opgeschoten gras (Ausden 1996).
5.7 Kruidenrijkdom en insectenaanbod voor kuikens
Volgens Oosterveld & Altenburg (2005) kan in het algemeen gesteld worden dat hoe kruidenrijker de vegetatie is,
des te meer ongewervelden aanwezig zijn in de vegetatie en op de bodemoppervlakte. Omdat gruttokuikens zich hoofdzakelijk voeden met in de vegetatie levende invertebraten (Schekkerman 1997), impliceert dit dat kruidenrijke vegetaties de beste foerageermogelijkheden bieden voor opgroeiende kuikens. Het is bekend dat bij bemesting de soortenrijkdom van vegetaties afneemt en niet bemeste percelen kruidenrijker zijn (o.a. Willems et al. 1993, Oosterveld & Altenburg 2005). In de niet bemeste percelen van Molenmeers lag de soortenrijkdom inderdaad veel hoger in vergelijking met bemeste percelen van Broekmeers (zie 4.6). Terwijl de proefvlakken van Broekmeers gedomineerd werden door grassen waren de proefvlakken van Molenmeers veel kruidenrijker. De vegetatie van Molenmeers zal bijgevolg veel geschikter zijn voor gruttokuikens.
Door te broeden op bemeste percelen die intensief bewerkt worden trappen grutto’s in een ecologische val. Niet alleen omdat er grotere verliezen optreden door een hogere intensiteit aan landbouwwerkzaamheden, maar ook omdat een hoge mestgift het voedsel voor de kuikens hypothekeert. Het grootste effect is afkomstig van het sneller maaien van bemeste percelen. Na de eerste snede op agrarische percelen worden dubbel zoveel invertebraten gevonden op nog niet gemaaide reservaatpercelen (Schekkerman 1997). Met deze reservaatpercelen worden weidevogelreservaten bedoeld, waar in Nederland een lichte bemesting met vaste mest toegepast wordt (streefdoel is daar 100 kg N ha-1 jaar-1, inclusief atmosferische depositie van ongeveer 50 kg N ha-1 jaar-1). Deze factor is voor weidevogels zeer relevant aangezien de eerste snede op agrarisch grasland dikwijls samenvalt met de periode waarin de meeste kuikens aanwezig zijn. Ook de biomassa van vegetatiebewonende arthropoden neemt af met toenemende bemesting, al treedt deze afname voornamelijk op onder de 100 kg N ha-1 jaar-1. Van zodra deze grens overschreden wordt, lijkt de biomassa niet meer verder te dalen (Siepel 1990).
Het totaal aantal of de biomassa arthropoden is volgens Kleijn et al. (2007) mogelijk geen goede maat voor de voedselbeschikbaarheid van gruttokuikens omdat het grootste deel hiervan bestaat uit prooidieren die te klein zijn om energetisch aantrekkelijk te zijn. Over het belang van de beschikbaarheid van grote prooidieren voor de ontwikkeling van gruttokuikens is echter nog weinig geweten en verder onderzoek is absoluut noodzakelijk (Kleijn et al. 2007). Aangezien de grootste insecten gevonden worden op niet bemeste percelen (Siepel 1990) zijn deze waarschijnlijk het beste biotoop voor gruttokuikens.
Bovendien scoren schrale, kruidenrijke vegetaties volgens Kleijn et al. (2007) ook het beste op vlak van fysieke geschiktheid voor gruttokuikens. De vegetatie blijft daar tijdens het hele voorjaar vrij ijl waardoor kuikens hier het minste energie zullen verspillen tijdens hun zoektocht naar prooien in de vegetatie. De vegetatie van percelen met een gangbare mestgift wordt meestal al te dicht in mei waardoor deze ongeschikt wordt voor foeragerende gruttokuikens. Om deze reden raden Kleijn et al. (2007) aan om uitgestelde maaidata vooral te plaatsen op kruidenrijke, lichter bemeste percelen of op percelen waar de gewasproductie door andere factoren, zoals een hoge waterstand, beperkt wordt. Wanneer beheerpakketten met uitgestelde maaidatum uitgevoerd worden op zwaar bemeste grond dragen deze niet bij tot het verbeteren van de voedselsituatie van gruttokuikens. Bijkomend stellen Kleijn et al. (2007) dat hergroeide percelen niet tot kuikenland kunnen gerekend worden omdat deze niet geschikt zijn voor kuikens. Na het maaien bieden ze zo goed als geen voedsel voor gruttokuikens en wanneer door hergroei het voedselaanbod stilaan verbetert is de vegetatie meestal al te dicht.
Als besluit kan gesteld worden dat schrale, kruidenrijke vegetaties zowel via het voedselaanbod als via de fysieke geschiktheid de grootste bijdrage zullen leveren tot het foerageersucces en de daarmee gepaard gaande overleving van gruttokuikens. Vegetaties onder natuurbeheer, zoals te vinden in Molenmeers, zijn dus ideaal kuikenland. Volgens Melman et al. (2008) moet minstens 1,4 ha geschikt kuikenland beschikbaar zijn per gruttokoppel. De sterker bemeste percelen van Broekmeers dragen minder bij tot het foerageersucces en de overleving van kuikens en zullen bijgevolg gemeden worden door gruttofamilies. Het noordelijk deel van Broekmeers is de laatste jaren wel natter geworden, waarschijnlijk door slecht onderhoud van bepaalde grachten. Door de vernatting en de daaraan gekoppelde daling van de mestgift is de vegetatie daar korter, kruidenrijker en dus geschikter geworden voor kuikens. Bovendien hebben grutto’s volgens Raes et al. (2009) bij vestiging in hun territoria een voorkeur voor kruidenrijke vegetaties. Dit kan ook verklaren waarom het leeuwendeel van de koppels in die overgangszone broedt en foerageert.
5.8 Een toekomst voor grutto’s? Aanbevelingen voor de inrichting en het beheer.
Uit deze studie blijkt dat de gruttodichtheid in de Kalkense Meersen nog zeer laag ligt in vergelijking met andere weidevogelgebieden in Nederland (Schekkerman et al. 2005). Het gebied bezit vermoedelijk wel het potentieel om een grotere gruttopopulatie uit te bouwen maar op dit moment wordt de geschiktheid van het habitat mogelijk verlaagd door tal van factoren zoals de beperkte openheid met de daaraan gekoppelde predatiedruk, verkeer, interacties tussen bemesting en grondwaterstand op het voedselaanbod voor volwassen vogels en kuikens,...
Op basis van literatuur en eigen bevindingen kan ik enkele maatregelen aanbevelen voor de toekomstige inrichting en beheer van de Kalkense Meersen. Deze maatregelen kunnen de populatiegrootte en het broedsucces van de grutto’s en andere weidevogels bevorderen.
De openheid van het gebied is zeer belangrijk. De predatiedruk neemt af met de openheid van het landschap. Kleine landschapselementen en bosjes bieden enerzijds beschutting en jachtmogelijkheden aan marterachtigen en vossen waardoor ze gemakkelijker ongezien bij de nesten van weidevogels geraken. Gevarieerde landschappen waarin een intense afwisseling optreedt in bodemgebruiksvormen (bossen, graslanden,..) en bijhorende lineaire structuren (perceelsranden, houtkanten,..) bieden hen ook meer mogelijkheden tot de uitbouw van grotere populaties (Verkem et al. 2003). Anderzijds fungeren bomen(rijen) en bosjes als nest- en uitkijkplaatsen voor gevleugelde predators zoals buizerd en zwarte kraai. Kraaiachtigen maken gebruik van zoekbeelden en leren snel waardoor weidevogels met zichtbare en bereikbare nesten gevoelig zijn (Bos & Vugteveen 2005). De verstoringsafstand van opgaande begroeiing en bos bedraagt respectievelijk 100 en 200 m (Oosterveld & Altenburg 2005). De bosjes, bomen(rijen) en overtollige weidepaaltjes worden dus best verwijderd. Dit geldt vooral voor Molenmeers, Langendonk en de randen van Broekmeers, maar ook voor de populierenbosjes op Belham en Scherenmeersen en de populieren langs de Belhamstraat. Broekmeers is nog een zeer open gebied wat waarschijnlijk één van de factoren is voor het hogere BTS dat daar gevonden werd. In een open landschap kunnen predators sneller opgemerkt en efficiënter verjaagd worden.
Een tweede maatregel die aangeraden wordt is het verhogen van de grondwatertafel. Een hogere watertafel heeft een negatieve invloed op grondpredatoren zoals zoogdieren (Teunissen et al. 2005). Bellebaum & Bock (2009), die de invloed van grondpredatoren en waterstand op het broedsucces van kieviten onderzochten, concluderen dat de overleving van kievitjongen stijgt wanneer water langer vastgehouden wordt in de jongenfase. Het inrichten van bredere watergangen waar zoogdieren moeilijk langs kunnen kan ook helpen. Om predatie tegen te gaan is het uitschakelen van één of enkele predatoren geen goed idee omdat andere predators de rol dan overnemen. Predatie (en verstoring) vormen vooral een probleem in combinatie met andere verliesoorzaken. Simulaties waarbij alle predatie uitgeschakeld werd boden nog geen garanties voor het voortbestaan van een populatie (Teunissen et al. 2005).
Verhogen van de watertafel heeft ook een indirecte invloed op de voedselbeschikbaarheid. Doordat een minder hoge en kruidenrijkere vegetatie gevormd wordt, zullen percelen geschikter worden als foerageergebied voor gruttokuikens. De voedselomstandigheden van de zeer korte vegetatie in de nabijheid van plasdras situaties zijn bovendien ideaal voor kieviten en tureluurs. De aanwezigheid van deze korte vegetatie zal bovendien de nood aan gemaaide percelen voor kieviten beperken. De plasjes zelf bieden tevens een alternatieve voedingsbron voor deze soorten, die zich ook voeden met aquatische larven van ongewervelden (Ausden 1996). Een hoge waterstand verlaat het tijdstip waarop de bodem ondoordringbaar wordt en regenwormen dieper in de bodem kruipen. Op deze manier blijft het voedselaanbod voor volwassen vogels langer op peil wat ook van belang kan zijn voor vervolglegsels (schekkerman 1997). Water tot in het maaiveld heeft echter ook negatieve gevolgen op het aantal wormen (Ausden et al. 2001). Om deze reden moeten ook iets drogere percelen aanwezig zijn in een weidevogelgebied. Siepel et al. (1990) en Schekkerman (1997) vonden zelfs dat iets drogere gronden meer macroinvertebraten herbergen dan natte. In de studie van Schekkerman (1997) was dit verschil echter niet significant.
Naast de invloed op de voedselbeschikbaarheid speelt de vochtigheid ook een rol in de geschiktheid van het nesthabitat. Grutto’s zijn niet zo kritisch maar soorten zoals kemphaan (Philomachus pugnax), watersnip, zomertaling en slobeend (Anas clypeata) vereisen water tot in het maaiveld voor hun vestiging in grasland. De korte vegetatie die zich ontwikkelt bij hoge waterstanden wordt geprefereerd door kieviten en tureluurs (Ausden 1996). Op minder natte plaatsen kan het gras eerder in het voorjaar opschieten wat voordeling is voor de grutto’s.
Een weidevogelgebied richt zich in de eerste plaats op zoveel mogelijk soorten. Om aan al de vereisten van zowel kuikens als volwassen vogels van verschillende soorten te voldoen wordt best een hydrologische mozaïek gecreëerd met vochtgehaltes die van zeer nat tot vochtig gaan. Zo’n mozaïek kan ontstaan door enerzijds de watertafel te verhogen en anderzijds microreliëf te creëren waardoor lage, zeer natte delen afgewisseld worden met iets hoger gelegen en dus drogere delen. In het algemeen kan de effectiviteit van agrarische natuurbeheersovereenkomsten bevorderd worden door het verhogen van de grondwatertafel. Bovendien wordt de densiteit en het aantal territoria voornamelijk verklaard door de grondwaterdiepte en het gemak waarmee voedsel gevonden wordt (Verhulst et al. 2007).
Behalve via de grondwaterstand kan de geschiktheid van het habitat voor weidevogels ook aangepast worden via het beheer. Wat de voedselvoorziening betreft, biedt natuurbeheer zonder bemesting en een uitgestelde maaidatum het beste kuikenland (Kleijn et al. 2007). Bij nog intensief bemeste percelen biedt een uitgestelde maaidatum geen meerwaarde voor gruttokuikens. Toch scoren reeds gemaaide percelen nog slechter (Kleijn et al. 2007). Kieviten zijn minder gebaat bij late maaidata en pas gemaaide percelen kunnen voor hen een meerwaarde bieden als geen korte vegetatie aanwezig is. Voorbeweide percelen zijn ook belangrijk als foerageergebied voor weidevogelkuikens. In de structuurrijke vegetatie en rond de uitwerpselen van koeien worden veel insecten gevonden, zeker voor kievitkuikens (Beintema et al. 1995). De bodemfauna wordt in het algemeen bevorderd door een lichte bemesting, behalve wanneer er uittredende kwel tot in het maaiveld aanwezig is. In de Kalkense Meersen is dat het geval waardoor het aan te raden is terughoudend te zijn met bemestingsplannen. Men onderzoekt beter eerst of nulbemesting wel tot een afname van de bodemfauna leidt. Als dit niet het geval is hypothekeert de bemesting alleen maar het voedsel voor de kuikens. Als toch gekozen wordt voor bemesting opteert men best voor vaste mest met een maximum hoeveelheid van 100 kg N ha-1 jaar-1 (inclusief jaarlijkse atmosferische depositie).
Variatie in vegetatiehoogte en geschiktheid als broedplaats kan eveneens geregeld worden via het beheer (naast de watertafel). Door een verlaagde mestgift zal de vegetatie weliswaar niet zo hoog opschieten, wat bevorderlijk is voor kieviten, maar niet voor grutto’s. Dit kan opgelost worden door nabegrazing. Op deze manier wordt structuur aangebracht waardoor in het volgende voorjaar snel pollen aanwezig zijn waarin grutto’s hun nest kunnen maken (Oosterveld & Altenburg 2005).
Verstoring kan tegengegaan worden door het bannen van verkeer en eventueel ook door het verbieden van zachte recreatie langs kerngebieden tijdens het broedseizoen (aangezien mensen dikwijls hun hond loslaten).
De verschillende factoren die bijdragen tot een lager broedsucces zoals predatie, verliezen door landbouwwerkzaamheden, slechte habitatkwaliteit en verstoring zijn meestal geen probleem op zichzelf. Het is de combinatie van al deze factoren die het de weidevogels tegenwoordig lastig maakt. Daarom moeten pogingen om het broedsucces en populaties van weidevogels te vergroten een holistische aanpak kennen waar al deze factoren tegelijk aangepakt worden. Voor de Kalkense Meersen bestaat de oplossing hoogstwaarschijnlijk in het creëren van een mozaïek in een meer open en rustiger gebied. Deze mozaïek bevat zowel een mozaïek in hydrologische omstandigheden als een mozaïek in maai- en beweidingsbeheer. Zo wordt een breed spectrum aan habitattypes beschikbaar en op deze manier is men in staat maximale ecologische potenties te creëren voor verschillende soorten weidevogels. Tenslotte is continuïteit in het beheer één van de belangrijkste aspecten omdat de broedplaatstrouw bij grutto’s gekoppeld is aan het broedsucces in de voorgaande jaren (Schekkerman et al. 2005).
Naast deze algemene richtlijnen, waarvan de meeste ook opgenomen zijn in de inrichtingsschets van het geactualiseerde Sigmaplan (zie Van Ryckegem et al. 2010), is het ook van belang te letten op ‘details’. Oevers van beken kuikenvriendelijk inrichten kan bijvoorbeeld ook vele kuikenlevens redden en biedt bovendien ook tal van foerageermogelijkheden.
De inrichting van het gebied en overgang naar verschillende habitattypes dient wel geleidelijk te gebeuren om ongewenste effecten tegen te gaan. Als men bijvoorbeeld in een korte periode overschakelt van zware bemesting op nulbemesting kan pitrus oprukken en dit is niet bevorderlijk voor weidevogels. Percelen die voor het eerst de winter zullen doorbrengen met water tot boven het maaiveld gaan best de winter in met een zo kort mogelijke vegetatie, om extreem anoxische condities te vermijden (Ausden et al. 2001). Zo kan mogelijk het negatieve effect van overstroming op de regenwormenstand beperkt worden. Via nabegrazing gaat de grasmat kort en structuurrijk de winter in.
Om de condities voor grutto’s in het Aubroek geschikter te maken gelden dezelfde richtlijnen als voor de Kalkense Meersen. Al is dit waarschijnlijk niet haalbaar op die schaal. Een verdere uitbouw van laat gemaaide percelen zou al een pluspunt zijn. Men moet wel beseffen dat het uitstellen van de maaidatum zonder extensiever beheer geen meerwaarde levert voor gruttokuikens (zie 5.7 en Kleijn et al. 2007).
Voor het absoluut kwantificeren van broedsucces in de toekomst beveel ik methodes aan die gebruik maken van zenderen en/of kleurringen (o.a. schekkerman et al. 2005) en dit in combinatie met de Mayfieldmethode (Mayfield 1975), waarmee dagelijkse overlevingskansen van eieren en jongen berekend worden. Enkel zo kunnen betrouwbare uitspraken gedaan worden over het aantal jongen dat koppels gemiddeld grootbrengen en of dit voldoende is om de populatie in stand te houden. Als er echter geen tijd en geld voorhanden is en men wenst enkel een indicatie te krijgen van het broedsucces dan volstaat een relatieve maat zoals het BTS (zie instructie alarmtellingen, Nijland & van Paassen 2007). Er moet echter wel voorzichtig omgesprongen worden met de interpretatie van de behaalde BTS-waarden als men te maken heeft met weinig broedparen en kleine gebieden, waar bovendien kans is op emi- en immigratie van gruttogezinnen. Als deze methode toegepast wordt zou ik, in tegenstelling tot wat ik gedaan heb, wel een vaste looproute volgen die door het grasland gaat. Zo zal een nauwkeurigere schatting verkregen worden van het BTS, zeker in grotere gebieden.
5.9 Conclusies
1. In de Kalkense Meersen kwamen in 2009 24 koppels grutto en minstens 55 koppels kievit tot broeden.
In het Aubroek bedroeg het aantal broedparen grutto en kievit respectievelijk 5 en 18.
2. Het weidevogelproject resulteert (voorlopig) nog niet in een toename van de gruttopopulatie in de Kalkense Meersen. Aanwas van gruttojongen die opgroeiden tijdens het project is pas te verwachten vanaf 2010, wat dus de nood aan inventarisatie in de toekomt benadrukt. Evaluaties op basis van aantallen zijn echter moeilijk te interpreteren. Daarom evalueert men beter het broedsucces of een relatieve maat ervan, zoals het bruto territoriaal succes (BTS).
3. Enkel in Broekmeers was het BTS hoog genoeg om de gruttopopulatie in stand te houden. Dit is een eerste indicatie die aangeeft dat het weidevogelproject resultaat oplevert. Het lage BTS in de reservaatzone is waarschijnlijk te wijten aan predatie en verstoring. In het Aubroek speelt naast deze factoren ook de vroege maaidatum mee.
De BTS-waarden moeten met voorzichtigheid geïnterpreteerd worden en dienen eerder ter vergelijking van de gebieden dan als exacte waarden voor het broedsucces.
4. Het weidevogelproject zorgt naast een reductie in legselverliezen effectief voor een hoger aandeel niet gemaaid grasland, wat de overleving van gruttokuikens bevordert. In vergelijking met gebieden zonder uitgestelde maaidata zal de grootte van het verschil afhankelijk zijn van het weer en de oppervlakte aan beheersovereenkomsten. Uitgestelde maaidata bieden geen meerwaarde voor gruttokuikens op intensief bemeste percelen. Daarom worden deze beheersovereenkomsten beter toegepast op kruidenrijke, lichter bemeste percelen of percelen waar de gewasproductie door andere factoren beperkt wordt.
5. De overleving van legsels en kuikens is afhankelijk van het weer. Het weer heeft namelijk een invloed op de maaidata en het foerageersucces van kuikens. Het weidevogelproject speelt via het uitstellen van de eerste snede in op deze weersinvloeden en het effect ervan zal maximaal zijn bij warm en droog weer. Het weer heeft bovendien een invloed op de voedselomstandigheden voor volwassen vogels.
6. De eerste broedende grutto’s van de Kalkense Meersen werden waargenomen in 1967. Nadien steeg de populatie tot een maximum van 30 koppels in 2002. De laatste jaren stagneert de populatie rond 25 koppels. Deze stagnatie is vermoedelijk te wijten aan een uitbreiding van de populatie naar nabijgelegen randgebieden als gevolg van een mindere geschiktheid van de Kalkense Meersen.
7. Tijdens deze populatiegroei zijn de kerngebieden sterk verschoven. Na Wijmeers werd de reservaatzone belangrijker maar de laatste jaren neemt Broekmeers deze rol over. De oorzaken van de verschillende verschuivingen zijn vermoedelijk te zoeken bij veranderingen in het beheer en verstoring onder de vorm van predatie, verkeer, landbouwwerkzaamheden of de aanwezigheid van andere organismen die het habitat ongeschikt maken.
8. Het voedselaanbod onder de vorm van regenwormen was groter in bemeste percelen van Broekmeers in vergelijking met niet bemeste percelen in Molenmeers. Het aantal en de biomassa regenwormen neemt meestal toe met bemesting maar er zijn verschillende interacties mogelijk met het weer, de grondwaterstand en het type bemesting. In gebieden met een invloed van kwel is men best terughoudend met bemesting.
9. De vegetatie was bij de start van het broedseizoen hoger in Broekmeers dan in Molenmeers. Aangezien grutto’s een hoge vegetatie als nestplaats verkiezen, is Broekmeers waarschijnlijk geschikter als nestplaats.
10. Kruidenrijke, schrale vegetaties staan bekend als het beste kuikenland, zowel qua voedselaanbod (insecten) als qua fysieke geschiktheid. Niet bemeste percelen in Molenmeers zijn soortenrijker en kruidenrijker als bemeste percelen in Broekmeers en daarom geschikter als kuikenland.
11. De bemonstering van regenwormen, vegetatiehoogte en de kruidenrijkdom in deze studie is niet representatief voor de totale oppervlakte van Broekmeers, maar enkel voor de iets drogere (lees: minder natte), meer bemeste percelen. De laatste jaren is het noordelijk deel van Broekmeers natter geworden met een lagere bemesting en een korte, kruidenrijke vegetatie als gevolg. Deze evolutie zorgt tegenwoordig voor een aanbod aan verschillende habitattypes tijdens het broedseizoen. Plasdras situaties in het voorjaar, een overgang van kort naar lang gras en van kruidenrijke naar lange, forse grassen, kortom een natuurlijke mozaïek. Als daarbij het weidevogelproject in rekening gebracht wordt, dat in 2010 bijna de hele oppervlakte van Broekmeers beslaat, maken deze omstandigheden Broekmeers tot een topgebied voor weidevogels! De verschuiving van de gruttokoppels naar Broekmeers is daarom vermoedelijk niet enkel te wijten aan aparte factoren zoals het aantal regenwormen of de langere vegetatie, maar aan het totale plaatje dat Broekmeers vandaag biedt.
12. Om een breed spectrum aan weidevogels een optimaal habitat te kunnen bieden, is het noodzakelijk de Kalkense Meersen volgens en holistische aanpak beter in te richten en (extensiever) te beheren. De ideale oplossing is het creëren van een mozaïek in een meer open, rustiger en natter gebied. Deze mozaïek bevat zowel een mozaïek in hydrologische omstandigheden als een mozaïek in maai- en beweidingsbeheer. Zo wordt een breed assortiment aan habitattypes beschikbaar en is men in staat maximale ecologische potenties te creëren voor verschillende soorten weidevogels.

Juichende grutto (Yves Adams © Vilda)
6 Samenvatting
Bijna alle broedende weidevogels in Europa vertonen de laatste decennia een afname in populatiegrootte.
De voornaamste oorzaken van deze achteruitgang zijn de intensivering van het agrarisch graslandgebruik en de toegenomen predatiedruk, met een daling in het broedsucces als gevolg. In Nederland worden weidevogels al redelijk lang beschermd via agrarisch natuurbeheer maar dit levert vooralsnog niet het gewenste resultaat. De beheersovereenkomsten bieden wel een hogere uitkomstkans van legsels maar de mortaliteit van kuikens ligt meestal nog te hoog.
Ondanks de negatieve vaststellingen in Nederland en andere landen inzake populatiegrootte, wordt deze negatieve trend niet vastgesteld in België. Er is zelfs sprake van een toename van het broedbestand bij kievit en grutto. Ook in het studiegebied van dit onderzoek, de Kalkense Meersen, steeg de gruttopopulatie tijdens de afgelopen jaren. Een specifiek weidevogelbeheer is nodig in het agrarische gebied om de weidvogels in stand te houden. Om deze reden en als voorbereiding op het geactualiseerde Sigmaplan, is het Regionaal Landschap Schelde-Durme in opdracht van het Agentschap voor Natuur en Bos en in samenwerking met de Provincie Oost-Vlaanderen in 2008 gestart met een soortenbeschermingsproject rond weidevogels in de Kalkense Meersen. Deze studie werd opgezet om na te gaan of de getroffen maatregelen effectief tot resultaten leiden en vormt dus een evaluatie van dit project, met een focus op de grutto.
Het oorspronkelijke opzet van het onderzoek streefde vier doelstellingen na. De eerste doelstelling was het bepalen van het aantal broedparen van grutto en kievit in de Kalkense Meersen en het Aubroek. De effectiviteit van het weidevogelproject werd enerzijds onderzocht door het bruto territoriaal succes (BTS) van de grutto’s te bepalen in drie deelgebieden. Anderzijds werd onderzocht of het weidevogelproject in vergelijking met gangbaar agrarisch grasland voor een hoger aandeel niet gemaaid grasland zorgt in de kuikenperiode. Om effecten van het weidevogelproject op de populatiegrootte te evalueren en om bepaalde trends waar te nemen en te verklaren, werd een historisch overzicht opgesteld van de gruttopopulatie en de broedlocaties. Uit dit historisch overzicht bleek een duidelijke verschuiving van broedlocaties naar bemeste percelen. Deze vaststelling riep nieuwe vragen op en om deze van antwoord te dienen werden nieuwe doelstellingen toegevoegd aan het onderzoek. Deze hadden als doel het verschil in aantal en biomassa regenwormen, vegetatiehoogte en kruidenrijkdom te bepalen tussen bemeste percelen in Broekmeers en niet bemeste percelen in Molenmeers.
In 2009 broedden 24 koppels grutto en minstens 55 koppels kievit in de Kalkense Meersen, in het Aubroek waren dit er respectievelijk 5 en 18. Daaruit blijkt dat het weidevogelproject (voorlopig) nog niet tot een toename van de gruttopopulatie leidt. Evaluaties op basis van populatiegrootte zijn echter moeilijk te interpreteren omdat de gevolgen van bv. emigratie, immigratie en mortaliteit moeilijk in te schatten zijn. Het is daarom beter om beschermingsmaatregelen te evalueren via het broedsucces of een relatieve maat ervan, zoals het BTS. De eerste indicatie voor de werking van het weidevogelproject komt van de BTS-waarden, die enkel in Broekmeers (90,9%) voldoende hoog waren om een populatie in stand te houden. De lage waarde in de reservaatzone (21,54%) is waarschijnlijk te wijten aan predatie en verstoring. In het Aubroek speelt naast deze factoren ook de vroege maaidatum mee. Het Aubroek haalde een BTS van 50%, maar deze waarde is niet representatief omwille van het lage aantal broedkoppels. Naast deze hoge BTS-waarde zorgt het weidevogelproject in Broekmeers effectief voor een hoger aandeel niet gemaaid grasland, in vergelijking met het Aubroek. In Broekmeers was tijdens de belangrijkste kuikenperiode (mei) steeds minstens 40% (en max. 77%) meer ongemaaid grasland aanwezig dan in het Aubroek. Dit zal ongetwijfeld bijgedragen hebben tot een hogere overleving van jongen aangezien deze positief gecorreleerd is met het aanbod niet gemaaid grasland. De overleving van gruttokuikens is bovendien afhankelijk van het weer. Het weidevogelproject speelt via het uitstellen van de eerste snede in op deze weersinvloeden en biedt naast een reductie in legselverliezen ook effectief een hoger aandeel niet gemaaid grasland aan.
Uit het historisch overzicht blijkt dat de eerst grutto’s zich vestigden in 1967. In de beginjaren werden maximum vier koppels waargenomen. Het is pas vanaf midden de jaren ’80 dat de populatie beginnen groeien is tot een maximum van 30 koppels in 2002. De laatste jaren stagneert de populatie rond 25 broedparen. Deze stagnatie is vermoedelijk te wijten aan een uitbreiding van de populatie naar nabijgelegen randgebieden. Als deze hypothese klopt, wijst dit op een mindere geschiktheid van de Kalkense Meersen. Deze mindere geschiktheid van sommige deelgebieden komt ook naar voor uit het historisch overzicht van de broedlocaties. Tijdens de populatiegroei zijn de kerngebieden van de grutto’s sterk verschoven. In de beginjaren broedden alle koppels in Wijmeers. Vanaf 1979 trad een verschuiving op naar de reservaatzone, dat het kerngebied bleef tot aan de piek van de populatiegroei in 2002. Na deze piek is Broekmeers aan een sterke uitbreiding begonnen en verloor Scherenmeersen al zijn koppels. Momenteel zijn Broekmeers, Langendonk en Belham de kerngebieden. De verschuivingen zijn vermoedelijk te wijten aan een verminderde geschiktheid van het habitat door veranderingen in het beheer en verstoring onder de vorm van predatie, verkeer, landbouwwerkzaamheden of de aanwezigheid van andere organismen die het habitat ongeschikt maken, zoals de uitheemse ganzen.
Het voedselaanbod onder de vorm van regenwormen was groter op bemeste percelen van Broekmeers in vergelijking met niet bemeste percelen van Molenmeers. In het algemeen is het aantal en de biomassa regenwormen positief gecorreleerd met de mestgift en de pH. Effecten van bemesting zijn dikwijls niet eenduidig vast te stellen door interacties met andere factoren zoals het weer, de grondwaterstand en het type bemesting. De beste resultaten worden bereikt met stalmest, maar in gebieden waar kwel tot in het maaiveld optreedt, moet men terughoudend zijn met bemesting. De zuurtegraad en de hoeveelheid organische stof blijkt daar van nature optimaal te zijn voor het in stand houden van goede regenwormpopulaties.
De vegetatie was bij de start van het broedseizoen hoger in Broekmeers dan in Molenmeers. Grutto’s verkiezen lang gras als nestplaats, wat Broekmeers geschikter maakt als nestgebied. De korte vegetatie van Molenmeers is dan weer ideaal voor kieviten, die liever hun nest in kort gras maken.
Niet bemeste percelen in Molenmeers zijn soorten- en kruidenrijker dan bemeste percelen in Broekmeers. Kruidenrijke, schrale vegetaties worden beschouwd als het beste kuikenland. Enerzijds omdat daar de grootste insecten aanwezig zijn, anderzijds omdat de vegetatie niet te dicht wordt, waardoor gruttokuikens minder energie verspillen tijdens hun zoektocht naar prooien.
Uit deze resultaten blijkt Broekmeers geschikter te zijn als nestplaats omwille van de hogere vegetatie en het hogere voedselaanbod voor volwassen grutto’s op het moment dat ze hun territoria kiezen. Nochtans biedt Molenmeers betere condities voor het opgroeien van de kuikens. Dit dilemma stelt zich tegenwoordig overal op gangbaar agrarisch grasland en grutto’s trappen in een ecologische val omdat ze een nestplaats kiezen die in principe minder geschikt is voor de overleving van hun jongen. Broekmeers is de laatste jaren echter niet meer te vergelijken met gangbaar agrarisch grasland omdat het noordelijk deel natter geworden is. Als gevolg zijn er in het voorjaar plasdras situaties aanwezig en overgangen van korte, kruidenrijke vegetaties naar lange, forse grasvegetaties. De ecologische val is op deze manier veel kleiner geworden en heeft van Broekmeers het beste weidevogelgebied in de Kalkense Meersen gemaakt.
Om de Kalkense Meersen geschikt te maken zodat verschillende weidevogels zich hier echt thuis voelen en een zichzelf regulerende populatie kunnen uitbouwen, moet het gebied volgens een holistische aanpak beter ingericht en (extensiever) beheerd worden. De ideale oplossing bestaat erin een mozaïek te creëren in een meer open, rustiger en natter gebied. Deze mozaïek bevat zowel een mozaïek in hydrologische omstandigheden als een mozaïek in maai- en beweidingsbeheer. Op deze manier zal een breed assortiment aan habitattypes beschikbaar zijn en is men in staat maximale ecologische potenties te creëren voor verschillende soorten weidevogels.
7 Dankwoord
Als eerste wil ik Peter Claus hartelijk bedanken. Hij heeft als begeleider in belangrijke mate bijgedragen tot dit werk door steeds tijd vrij te maken voor mij, via het aanbrengen van nuttige ideeën en commentaar, hulp bij het veldwerk, aanbieden van informatie en zoveel meer. Ik heb bovendien zeer veel bijgeleerd van hem. Daarnaast wil ik mijn promotor Johan Mertens en begeleidster Jenny De Laet bedanken voor de verdere ondersteuning en adviezen.
Roeland Cortois, Tomas Willems, Hans Matheve, Lennert Schepers en Peter Claus leverden een belangrijke bijdrage aan het veldwerk. Roeland Cortois en Martijn Vandegehuchte ben ik dankbaar voor de ondersteuning bij de statistische analyses. Landbouwer Emiel Van Petegem en Natuurpunt gaven toestemming om het regenwormenonderzoek op hun terreinen uit te voeren.
Veel van de gegevens die gebruikt werden voor het opstellen van het historisch verloop van de gruttopopulatie werden ter beschikking gesteld door Gunter De Smet, Rudi Van Onderbergen, Dirk De Mesel, Antoon Blondeel, Rudi Clinckspoor en Peter Claus.
Nuttig commentaar op eerdere versies van dit werk werd geleverd door Peter Claus, Jenny De Laet, Dominiek Decleyre en Hilde Hoogewijs.
Toestemming voor het gebruik van enkele foto’s werd verleend door Rollin Verlinde (Vilda) en Marc Van den Bril.
Dit onderzoek werd tenslotte mogelijk gemaakt door de Universiteit Gent en Regionaal Landschap Schelde-Durme.
8 Referenties
Altenburg, W. & Wymenga, E. (2000) Help, de grutto verdwijnt! De Levende Natuur, 101, 62-64.
Anderson, M.J., Gorley, R.N. & Clarke, K.R. (2008) PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods.
Ausden, M. (1996) The effects of raised water levels on food supply for breeding waders on lowland wet grassland. PhD Thesis, University of East Anglia, Norwich, UK.
Ausden, M., Sutherland, W.J. & James, R. (2001) The effects of flooding lowland wet grassland on soil macroinvertebrate prey of breeding wading birds. Journal of Applied Ecology, 38, 320-338.
Baines, D. (1990) The Roles of Predation, Food and Agricultural Practice in Determining the Breeding Success of the Lapwing (Vanellus Vanellus) on Upland Grasslands. Journal of Animal Ecology, 59, 915-929.
Beintema, A.J., Dunn, E. & Stroud, D. (1997) Birds and wet grasslands. Farming and birds in Europe: the Common Agricultural Policy and its Implications for Bird Conservation (eds Pain, D.J. & Pienkowski, M.D.), pp. 269-296. Academic Press, San Diego.
Beintema, A.J., Moedt, O. & Ellinger, D. (1995) Ecologische atlas van de Nederlandse weidevogels. Schuyt & Co, Haarlem.
Bellebaum, J. & Bock, C. (2009) Influence of ground predators and water levels on Lapwing Vanellus vanellus breeding success in two continental wetlands. Journal of Ornithology, 150, 221-230.
Besbeas, P., Freeman, S. N., Morgan, B. J. T. & Catchpole, E. A. (2002) Integrating mark-recapture-recovery and census data to estimate animal abundance and demographic parameters. Biometrics, 58, 540-547.
Borek, V., Morra, M.J., Brown, P.D. & Mccaffrey, J.P. (1995) Transformation of the Glucosinolate-Derived Allelochemicals Allyl Isothiocyanate and Allylnitrile in Soil. Journal of Agricultural and Food Chemistry, 43, 1935-1940.
Bos, F. & Vugteveen, P. (2005) Kraaiachtigen een bedreiging voor weidevogels? Een literatuuronderzoek naar de rol van kraaiachtigen als predator en de invloed daarvan op weidevogels. Wetenschapswinkel Biologie, Rapport 67. Rijksuniversiteit Groningen, Haren.
Brandsma, O.H. (1999) Het belang van bemesting voor het voedselaanbod van weidevogels. De Levende Natuur, 100, 118-123.
Busche, G. (1994) The Decline of Wet-Meadow Birds in Schleswig-Holstein Germany from 1950 to 1992. Journal Fur Ornithologie, 135, 167-177.
Caekebeke, L. & de Bruycker, P (1984) Het ornitologisch belang van de Kalkense Meersen. Wielewaal, 50, 338-356.
Claus, P. (2005) De broedvogels van de Kalkense Meersen. Tronk, 3, 8-25.
Čoja, T., Zehetner, K., Bruckner, A., Watzinger, A. & Meyer, E. (2008) Efficacy and side effects of five sampling methods for soil earthworms (Annelida, Lumbricidae). Ecotoxicology and Environmental Safety, 71, 552-565.
Curry, J.P. (2004) Factors affecting the abundance of earthworms in soils. Earthworm Ecology (ed C. A. Edwards), pp. 91-114. CRC press LLC, Boca Raton, FL.
Darwin, C. (1881) The Formation of Vegetable Mould Through the Action of Worms. Murray, London, UK.
Degezelle, T., Kongs, T., Martens, L., Vercoutere, B. & Hoffmann, M. (2004) Ontwerp- ecosysteemvisie Kalkense meersen en Berlare broek: een verkenning van natuurpotenties. AMINAL, Brussel.
Devos, K., Vermeersch, G. & Jacob, J.P. (2003) Population estimates of breeding waders in Belgium. Wader Study Group Bull.
Estevez, B., N'Dayegamiye, A. & Coderre, D. (1996) The effect on earthworm abundance and selected soil properties after 14 years of solid cattle manure and NPKMg fertilizer application. Canadian Journal of Soil Science, 76, 351-355.
Green, R.E. (1988) Effects of Environmental factors on the timing and succes of breeding of Common Snipe Gallinago gallinago (Aves, Scolopacidae). Journal of Applied Ecology, 25, 79-93.
Groen, N.M. (1993) Breeding site tenacity and natal philopatry in the black-tailed godwit Limosa l. limosa. Ardea, 81, 107-113.
Hagemeijer, W.J.M., Blair, M.J., van Turnhout, C., Bekhuis, J. & Bijlsma, R. (1997) EBCC Atlas of European Breeding Birds: their Distribution and Abundance. Poyser, London.
Hansen, S. & Engelstad, F. (1999) Earthworm populations in a cool and wet district as affected by tractor traffic and fertilisation. Applied Soil Ecology, 13, 237-250.
Huijsmans, J.F.M., Schröder, J.J., Vermeulen, G.D., de Goede, R. G. M., Kleijn, D. & Teunissen, W.A. (2008) Emissiearme mesttoediening. Ammoniakemissie, mestbenutting en nevenaspecten. Rapport 195. Plant Research International B.V., Wageningen.
Hut, H. & Helmig, F. (2003) Valt hier nog wat te vreten?: onderzoek naar de relaties tussen voedsel, zuurgraad en broedende weidevogels. Staatsbosbeheer, Regio Fryslân/Groningen-Drenthe.
Kleijn, D., Berendse, F., Smit, R. & Gilissen, N. (2001) Agri-environment schemes do not effectively protect biodiversity in Dutch agricultural landscapes. Nature, 413, 723-725.
Kleijn, D., Dimmers, W., van Kats, R., Melman, D. & Schekkerman, H. (2007) De voedselsituatie voor gruttokuikens bij agrarisch mozaïekbeheer. Alterra-rapport 1487. Alterra, Wageningen.
Kruk, M., Noordervliet, M.A.W. & terKeurs, W.J. (1997) Survival of black-tailed godwit chicks Limosa limosa in intensively exploited grassland areas in The Netherlands. Biological Conservation, 80, 127-133.
Kuijper, D.P.J., Wymenga, E., Van Der Kamp, J. & Tanger, D. (2006) Wintering areas and spring migration of the Black-tailes Godwit. Bottlenecks and protection along the migration route. A&W-rapport 820. Altenburg & Wymenga ecologisch onderzoek, Veenwouden.
Ma, W.C., Brussaard, L. & Deridder, J.A. (1990) Long-Term Effects of Nitrogenous Fertilizers on Grassland Earthworms (Oligochaeta, Lumbricidae) - Their Relation to Soil Acidification. Agriculture Ecosystems & Environment, 30, 71-80.
Mather, J.G. & Christensen, O. (1988) Surface movements of earthworms in agricultural land. Pedobiologia, 32, 399-405.
Mather, J.G. & Christensen, O. (1992) Surface Migration of Earthworms in Grassland. Pedobiologia, 36, 51-57.
Mayfield, H.F. (1975) Suggestions for calculating nest succes. Wilson Bull., 87, 456-467.
Melman, D., van de Boel, P., Dijkstra, J., Oosterveld, E., van Paassen, A., Schotman, A., Terwan, P. & van 't veer, R. (2008) Voorstel bouwstenen nieuwe weidevogelpakketten agrarisch natuurbeheer in een notendop. wat regelen we in Nederland, wat in Brussel? Rapport DK nr. 2008/098. Directie Kennis, Ministerie van LNV, Ede.
Nijland, F. (2002) Project alarm, een verkennend onderzoek naar territoriaal succes van scholekster, kievit, grutto en tureluur in de periode 1997-2000 in fyslan. Publcatie nr. 12. Bureau N, Leeuwarden.
Nijland, F., Schekkerman, H. & Teunissen, W. (in voorbereiding) Methodes monitoring weidevogels.
Nijland, F. & van Paassen, A. (2007) Instructie Alarmtellingen; tellingen van paren en gezinnen van scholekster, kievit, grutto, tureluur en wulp. Publicatie Bureau N nr. 27. Landschapsbeheer Nederland, Utrecht. , Leeuwarden.
Oosterveld, E. (2006) Betekenis van waterpeil en bemesting voor weidevogels. De Levende Natuur, 107, 134-137.
Oosterveld, E.B. & Altenburg, W. (2005) Kwaliteitscriteria voor weidevogelgebieden, met toetslijst. A&W-rapport 412. Altenburg & Wymenga ecologisch onderzoek, Veenwouden.
Ottvall, R. (2005) Breeding success and adult survival of Redshank Tringa totanus on coastal meadows in SE Sweden. Ardea, 93, 225-236.
Peach, W.J., Thompson, P. S. & Coulson, J. C. (1994) Annual and Long-Term Variation in the Survival Rates of British Lapwings Vanellus-Vanellus. Journal of Animal Ecology, 63, 60-70.
Pelosi, C., Bertrand, M., Capowiez, Y., Boizard, H. & Roger-Estrade, J. (2009) Earthworm collection from agricultural fields: Comparisons of selected expellants in presence/absence of hand-sorting. European Journal of Soil Biology, 45, 176-183.
Plum, N.M. & Filser, J. (2005) Floods and drought: Response of earthworms and potworms (Oligochaeta : Lumbricidae, Enchytraeidae) to hydrological extremes in wet grassland. Pedobiologia, 49, 443-453.
Raes, N., Hoogeboom, D., Baas, T., Visbeen, F. & Scharringa, K (2010) Grutto's in de Krommenieër Woudpolder, Weijenbus en Vroonmeer, Polder de Zeevang-West, en Waerland-Oost in 2009. Rapportnummer: LNHOA-010-03.
Schekkerman, H. (1997) Graslandbeheer en groeimogelijkheden voor weidevogelkuikens. IBN-rapport 292. IBN-DLO, Wageningen.
Schekkerman, H. (2008) Precocial problems. Shorebird chick performance in relation to weather, farming and predation. Proefschrift ter verkrijging van het doctoraat in de Wiskunde en Natuurwetenschappen aan de Rijksuniversiteit Groningen.
Schekkerman, H. & Beintema, A.J. (2007) Abundance of invertebrates and foraging success of Black-tailed Godwit Limosa limosa chicks in relation to agricultural grassland management. Ardea, 95, 39-54.
Schekkerman, H. & Müskens, G. (2000) Produceren grutto's Limosa limosa in agrarisch grasland voldoende jongen voor een duurzame populatie? Limosa, 73, 121-134.
Schekkerman, H., Teunissen, W. & Oosterveld, E. (2005) Resultaatonderzoek Nederland Gruttoland; broedsucces van grutto's in beheersmozaïeken in vergelijking met gangbaar agrarisch graslandgebruik. Alterra-rapport 1291. Alterra, Wageningen.
Schekkerman, H., Teunissen, W. & Oosterveld, E. (2008) The effect of 'mosaic management' on the demography of black-tailed godwit Limosa limosa on farmland. Journal of Applied Ecology, 45, 1067-1075.
Siepel, H. (1990) The influence of management on food size in the menu of insectivorous animals. Proceedings in experimental & applied Entomology, N.E.V. Amsterdam, 1, 69-74.
Siepel, H., Slim, P.A., Ma, W., Meijer, J., Wijnhoven, H.A.H, Bodt, J. & van Os, L.J. (1990) Effecten van verschillen in mestsoort en waterstand op vegetatie en fauna van klei-op-veen graslanden in de Alblasserwaard. RIN-rapport 90/8, pp. 49 p. RIN, Arnhem.
Standen, V. (1984) Production and Diversity of Enchytreids, Earthworms and Plants in Fertilized Hay Meadow Plots. Journal of Applied Ecology, 21, 293-312.
Stephens, P.A., Sutherland, W. J. & Freckleton, R. P. (1999) What is the Allee effect? Oikos, 87, 185-190.
Svensson, L., Grant, P.J., Mullarney, K. & Zetterström, D. (2008) Vogelgids van Europa. ANWB, Den Haag.
Teunissen, W., Schekkerman, H., Willems, F. & Majoor, F. (2008) Identifying predators of eggs and chicks of Lapwing Vanellus vanellus and Black-tailed Godwit Limosa limosa in the Netherlands and the importance of predation on wader reproductive output. Ibis, 150, 74-85.
Teunissen, W.A. (2000) Grutto alarm. SOVON-nieuws, pp. 13:14.
Teunissen, W.A., Schekkerman, H. & Willems, F. (2005) Predatie bij weidevogels. Op zoek naar de mogelijke effecten van predatie op de weidevogelstand. Soven-onderzoeksrapport 2005/11. Sovon Vogelonderzoek Nederland, Beek-Ubbergen.
Thorup, O. (2006) Breeding Waders in Europe 2000. International Wader Studies. International Wader Study Group, UK.
Timmerman, A., Bos, D., Ouwehand, J. & de Goede, R.G.M. (2006) Long-term effects of fertilisation regime on earthworm abundance in a semi-natural grassland area. Pedobiologia, 50, 427-432.
Trolliet, B. (2000) Plan de Gestion de l'Union Européenne pour le Vanneau huppé Vanellus vanellus. Contrat 97/162/3040/DEB/D2. Commission Européenne Direction générale XI. , Brussel.
van Damme, D. & Lostrie, C. (1987) Het kasteel en de meersen. Uitgeverij Groeninghe n.v.
Van Impe, J. (2003) Voortplantingssucces van kievit Vanellus vanellus, grutto Limosa limosa en tureluur Tringa totanus te Antwerpen-Linkeroever. Natuur.oriolus, 69, 45-59.
Van Ryckegem, G., Piesschaert, F. & Van den Bergh, E. (2010) Ecosysteemvisie cluster Kalkense meersen (zone 1). Studie t.b.v. aanleg overstromingsgebieden en natuurgebieden i.h.k.v. het SIGMAPLAN. INBO.R.2010.3.
Verhulst, J., Kleijn, D. & Berendse, F. (2007) Direct and indirect effects of the most widely implemented Dutch agri-environment schemes on breeding waders. Journal of Applied Ecology, 44, 70-80.
Verkem, S., De Maeseneer, J, Vandendriessche, B, Berbeylen, G & Yskout, S (2003) Zoogdieren in Vlaanderen. Ecologie en verspreiding van 1987 tot 2002. Natuurpunt Studie & JNM-Zoogdierenwerkgroep, Mechelen & Gent, België.
Vermeersch, G., Devos, K. & Anselin, A. (2006) Weidevogels in Vlaanderen. De Levende Natuur, 107, 75-80.
Vickery, J.A., Tallowin, J.R., Feber, R.E., Asteraki, E.J., Atkinson, P.W., Fuller, R.J. & Brown, V.K. (2001) The management of lowland neutral grasslands in Britain: effects of agricultural practices on birds and their food resources. Journal of Applied Ecology, 38, 647-664.
Visser, G.H. & Ricklefs, R.E. (1993) Development of Temperature Regulation in Shorebirds. Physiological Zoology, 66, 771-792.
Willems, J.H., Peet, R. K. & Bik, L. (1993) Changes in chalk-grassland structure and species richness resulting from selective nutrient additions. Journal of Vegetation Science, 4, 203-212.
Wilson, A.M., Ausden, M. & Milsom, T.P. (2004) Changes in breeding wader populations on lowland wet grasslands in England and Wales: causes and potential solutions. Ibis, 146, 32-40.
Wymenga, E. & Alma, R. (1998) Onderzoek naar de achteruitgang van weidevogels in het natuurreservaat De Gouden Bodem. A&W-rapport 170. Altenburg & Wymenga ecologisch onderzoek, Veenwouden.
Zhang, Y.S. & Talalay, P. (1994) Anticarcinogenic Activities of Organic Isothiocyanates - Chemistry and Mechanisms. Cancer Research, 54, S1976-S1981.
Internetbronnen :
www.kmi.be
www.gisoost.be
www.waarnemingen.be
9 Bijlagen
Bijlage I. Broedparen en bruto territoriaal succes
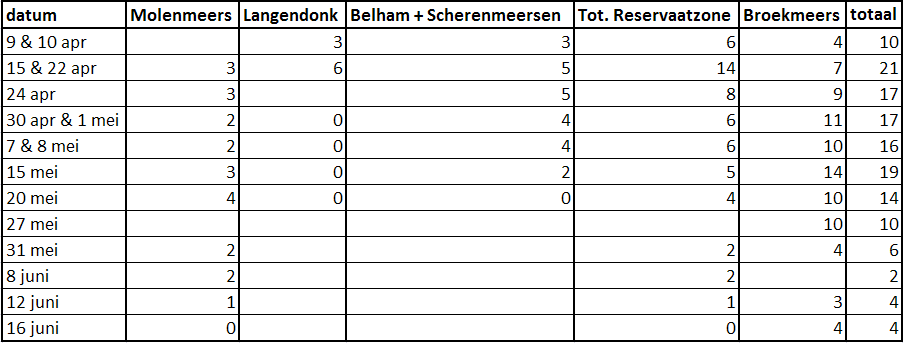
Tabel 9.1. Aantal broedparen dat op de aangegeven data (2009) werd vastgesteld in de verschillende deelgebieden. Aangezien het volledige studiegebied soms niet in één dag geïnventariseerd kon worden, zijn af en toe twee data samen genomen. Op die twee dagen werd het hele studiegebied geïnventariseerd. Een leeg vak duidt erop dat het deelgebied op die datum niet geteld werd.
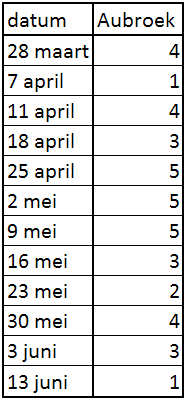
Tabel 9.2. Aantal broedparen dat op de aangegeven data (2009) werd vastgesteld in het Aubroek.
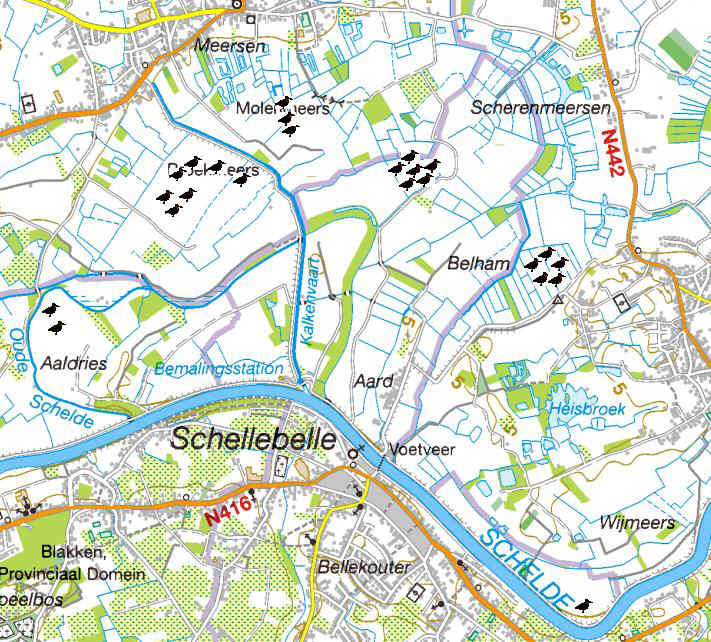
Figuur 9.1. Exacte locatie van de gruttonesten in 2009.
Bijlage II. Historisch verloop gruttopopulatie (randgebieden)
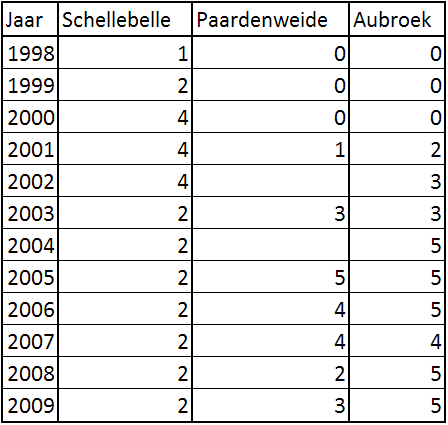
Tabel 9.3. Aantal broedparen in de randgebieden (1998-2009). Lege vakken verwijzen naar ontbrekende gegevens.
Bijlage III. Kruidenrijkdom

Tabel 9.4. Lijst met gedetermineerde plantensoorten op bemeste percelen in Broekmeers en niet bemeste percelen in Molenmeers.
| masterproef_de_grutto_in_de_kalkense_meersen_schepersrobbert.pdf |